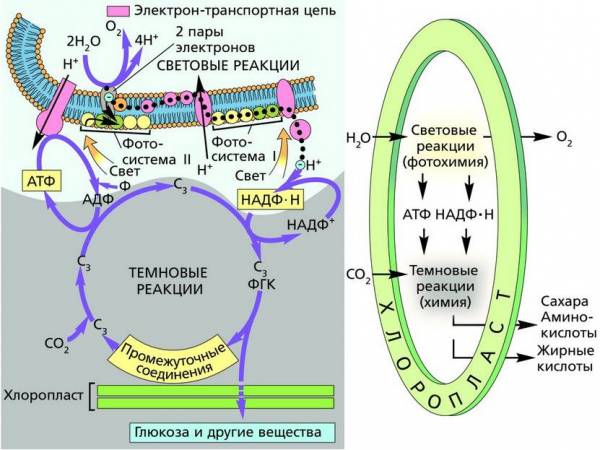
Фотосинтез – это сложный процесс, происходящий на свету в хлоропластах растительной клетки.
В процессе фотосинтеза различают два цикла реакций – две его фазы, последовательно и непрерывно идущие друг за другом, – световую и темновую.
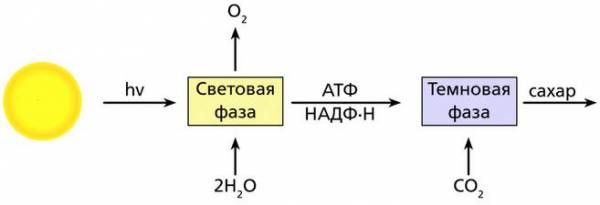 Схема фотосинтеза
Схема фотосинтеза
Рассмотрим сначала особенности световой фазы фотосинтеза.
Световая фаза фотосинтеза характеризуется тем, что здесь все процессы происходят только при участии энергии света. В основе световой фазы лежит функционирование двух фотосинтетических молекулярных структур – фотосистемы I и фотосистемы II.
Фотосистемы содержат по 250–400 молекул различных пигментов и белки. Все молекулы фотосистем способны поглощать свет, но только одна– основная молекула хлорофилла, являющаяся реакционным центром (РЦ), использует поглощенную энергию в фотохимических реакциях. Все остальные молекулы – антенные. В реакционных центрах сосредоточена лишь небольшая (1 %) часть хлорофилла, непосредственно участвующего в преобразовании энергии поглощенных фотонов в энергию химических связей. Различают фотосистемы по характеру поглощения света хлорофиллом реакционного центра. Максимум поглощения света фотосистемы I (ФС I) соответствует длине волны 700 нм (П700), антенных пигментов ФС I – хлорофиллов a (675 – 695 нм). Максимум поглощения фотосистемы II (ФС II) – 680 нм (П680), ее антенных пигментов – хлорофиллов a (670–683 нм). Электроны хлорофиллов последовательно возбуждаются в обеих фотосистемах.
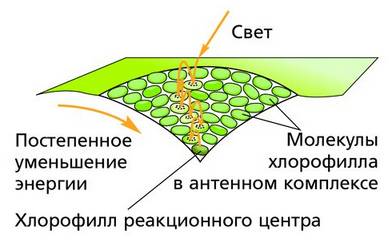 Антенный комплекс на тилакоидной мембране Фотосистема II. В первой фотосистеме (по историческим причинам получившей название фотосистемы II, или ФС II) в результате поглощения кванта света электроны хлорофилла a680 (П680) реакционного центра возбуждаются и поднимаются на более высокий энергетический уровень. Здесь они сразу же захватываются первым акцептором – белком феофитином (Фф). Получив электроны, феофитин становится сильным донором и сразу же отдает их другому акцептору, тот передает третьему и т. д. до тех пор, пока электроны не достигнут конечного акцептора, которым служит фотосистема I (ФС I). Цепь электронного транспорта, соединяющая две фотосистемы, в качестве переносчиков электронов включает в себя пластохинон (ПХ, PQ), отдельный электрон-транспортный комплекс цитохромов (так называемый b/f-комплекс) и водорастворимый белок пластоцианин (ПЦ, PC). Фотосистема I, получив электроны, тоже становится сильным донором, и ее электроны захватываются следующим акцептором – белком ферредоксином (Фд), а от него переходит к белку-ферменту ферредоксин-НАДФ-редуктазе, где используются на восстановление НАДФ + до НАДФ·Н. На рисунке показана цепь переноса электронов в фотохимических реакциях от ФС II к ФС I, которую часто называют Z-схемой. Стрелками показан поток электронов.
Антенный комплекс на тилакоидной мембране Фотосистема II. В первой фотосистеме (по историческим причинам получившей название фотосистемы II, или ФС II) в результате поглощения кванта света электроны хлорофилла a680 (П680) реакционного центра возбуждаются и поднимаются на более высокий энергетический уровень. Здесь они сразу же захватываются первым акцептором – белком феофитином (Фф). Получив электроны, феофитин становится сильным донором и сразу же отдает их другому акцептору, тот передает третьему и т. д. до тех пор, пока электроны не достигнут конечного акцептора, которым служит фотосистема I (ФС I). Цепь электронного транспорта, соединяющая две фотосистемы, в качестве переносчиков электронов включает в себя пластохинон (ПХ, PQ), отдельный электрон-транспортный комплекс цитохромов (так называемый b/f-комплекс) и водорастворимый белок пластоцианин (ПЦ, PC). Фотосистема I, получив электроны, тоже становится сильным донором, и ее электроны захватываются следующим акцептором – белком ферредоксином (Фд), а от него переходит к белку-ферменту ферредоксин-НАДФ-редуктазе, где используются на восстановление НАДФ + до НАДФ·Н. На рисунке показана цепь переноса электронов в фотохимических реакциях от ФС II к ФС I, которую часто называют Z-схемой. Стрелками показан поток электронов.
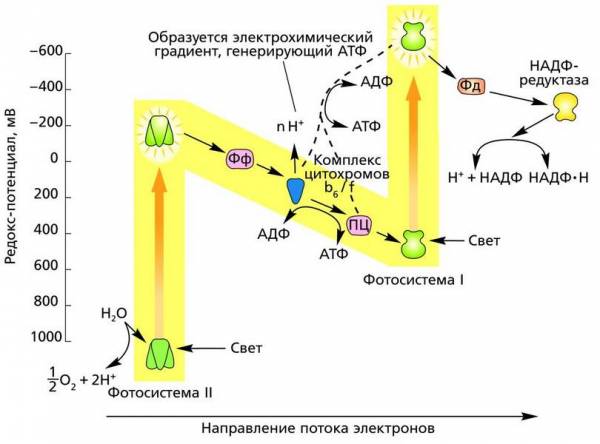 Z-схема переноса электронов (нециклический поток) в фотохимических реакциях фотосинтеза. Пунктиром показан циклический поток электронов
Z-схема переноса электронов (нециклический поток) в фотохимических реакциях фотосинтеза. Пунктиром показан циклический поток электронов
Сам перенос электронов по линейной цепи представлен на рисунке ниже.
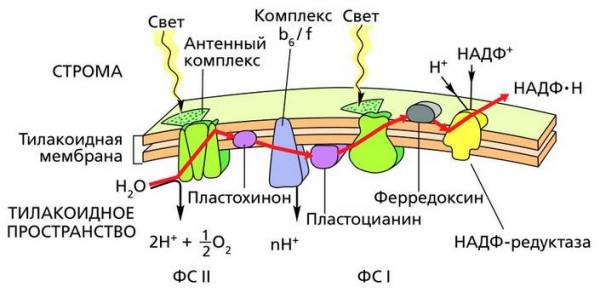 Электрон-транспортная цепь в мембране тилакоидов
Электрон-транспортная цепь в мембране тилакоидов
Следует отметить, что первым донором электронов для закрытия «дырки» в реакционном центре ФС II служит вода. Под действием света и в присутствии окислителей в хлоропластах происходит процесс разложения воды – фотолиз. Установлено, что фотолиз идет при участии вещества-переносчика – пластохинона и катализируется ферментами в присутствии ионов марганца, хлора и кальция. Весь комплекс соединений, обеспечивающих фотолиз, часто называют кислородвыделяющим комплексом. В итоге расщепления одной молекулы воды образуются 2 протона, 2 электрона и ½ молекулы кислорода. Процесс фотолиза воды можно выразить уравнением:
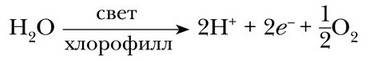 Фотолиз воды
Фотолиз воды
Образовавшийся свободный кислород как побочный продукт фотосинтеза диффундирует в окружающую среду или расходуется растительными клетками для дыхания.
В итоге в фотосистеме II по электрон-транспортной цепи переносятся электроны, полученные и от окисления воды, и от возбужденного светом хлорофилла а680 (П680).
Таким образом, световой этап фотосинтеза в ФС II и ФС I условно можно разделить на две стадии. Первая включает первичные световые реакции, обеспечивающие процессы активации электронов и их переноса в реакционных центрах. На второй световой стадии фотосинтеза происходят биохимические процессы, в которых образуется сильный восстановитель НАДФ·Н, синтезируется АТФ и при фотоокислении воды выделяется O2.
Транспорт электронов и все реакции второй световой стадии происходят в тилакоидной мембране хлоропластов. Ключевую роль в процессах трансформации световой энергии в хлоропластах и ее запасания в форме макроэргических связей АТФ играет АТФ-синтаза.
Световые реакции фотосинтеза
На пути продвижения электронов по электрон-транспортной цепи от ФС II к ФС I между акцепторами – пластохиноном и пластоцианином расположен комплекс цитохромов (b6/f), сопрягающий транспорт электронов с формированием электрохимического потенциала на мембране тилакоида. После окисления первого акцептора– пластохинона (ПХ) один из двух электронов, следуя по цепи через цитохром f – пластоцианин (ПЦ), направляется в фотосистему I, а второй снова поступает в комплекс цитохромов (b6/f) и, взаимодействуя с акцептором ПХ, осуществляет подряд два окисления. Таким образом происходит увеличение количества электронов, направляющихся по электрон-транспортной цепи к ФС I. Два следующих друг за другом окисления в комплексе цитохромов (b6/f) приводят к увеличению количества протонов, образующихся в этих реакциях. В итоге в тилакоидном пространстве накапливаются протоны, поступающие сюда и от фотолиза воды, и от комплекса цитохромов.
Накопление протонов в тилакоидном пространстве приводит к созданию в протонном резервуаре электрохимического градиента. Когда он достигает величины приблизительно 200 mW, протоны с внутренней стороны мембраны переносятся по электрохимическому градиенту на внешнюю через канал, образованный ферментом АТФ-синтазой (АТФазой). При этом происходит процесс фосфорилирования (образования АТФ). Поток протонов, поступающих через канал, служит источником энергии для синтеза АТФ, а перенесенный протон, взаимодействуя с электроном и молекулой окисленного НАДФ + , восстанавливает его до НАДФ·Н.
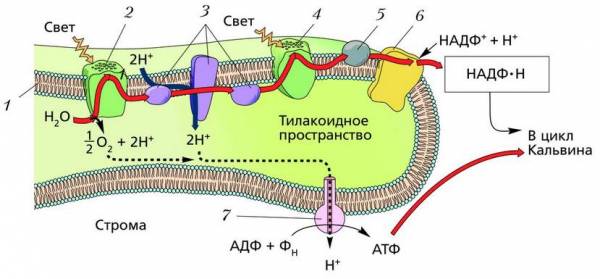 Механизм фотофосфорилирования в мембране тилакоида: 1 – внутренняя мембрана тилакоида; 2 – ФС II; 3 – переносчики электронов (ПХ, комплекс b6/f, ПЦ); 4 – ФС I; 5 – ферредоксин; 6 – НАДФ-редуктаза; 7 – АТФ-синтаза
Механизм фотофосфорилирования в мембране тилакоида: 1 – внутренняя мембрана тилакоида; 2 – ФС II; 3 – переносчики электронов (ПХ, комплекс b6/f, ПЦ); 4 – ФС I; 5 – ферредоксин; 6 – НАДФ-редуктаза; 7 – АТФ-синтаза
АТФ-синтаза – это целостный комплекс белковых молекул (субъединиц). В ходе катализа его (субъединица работает как стержень – ротор «молекулярного мотора», который попеременно вращается относительно неподвижных α- и β-субъединиц, – то против β-, то против α-субъединицы. Энергия конформационных изменений молекулярного комплекса трансформируется в энергию фосфатной связи АТФ. Модель АТФ-синтазы была предложена американским химиком Полом Бойером. За это открытие П. Бойер и английский химик Джон Уокер в 1997 году были удостоены Нобелевской премии.
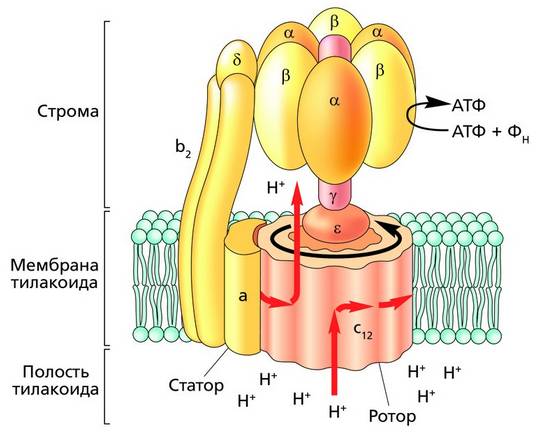 Строение АТФ-синтазы: a, b2, α, β, δ – закрепленные субъединицы; c12, ε, γ – вращающиеся субъединицы Фотосистема I. В реакционном центре фотосистемы I при возбуждении молекулы хлорофилла a700 (П700) энергией, равной двум квантам длинноволнового красного света, от нее «отрывается» пара электронов, которые переходят на более высокие вакантные орбитали и быстро захватываются акцепторами. Первым акцептором электронов выступает молекула хлорофилла a (A0) и хлорофилла a695 (A1). Далее следует серия переносчиков (железосерные белки), через которые электроны поступают на ферредоксин – сильный биологический восстановитель. От ферредоксина через фермент флавопротеин-ферредоксин-НАДФ-редуктазу электроны идут на восстановление НАДФ + до НАДФ·Н.
Строение АТФ-синтазы: a, b2, α, β, δ – закрепленные субъединицы; c12, ε, γ – вращающиеся субъединицы Фотосистема I. В реакционном центре фотосистемы I при возбуждении молекулы хлорофилла a700 (П700) энергией, равной двум квантам длинноволнового красного света, от нее «отрывается» пара электронов, которые переходят на более высокие вакантные орбитали и быстро захватываются акцепторами. Первым акцептором электронов выступает молекула хлорофилла a (A0) и хлорофилла a695 (A1). Далее следует серия переносчиков (железосерные белки), через которые электроны поступают на ферредоксин – сильный биологический восстановитель. От ферредоксина через фермент флавопротеин-ферредоксин-НАДФ-редуктазу электроны идут на восстановление НАДФ + до НАДФ·Н.
НАДФ·Н (никотинамидадениндинуклеотидфосфат восстановленный) – кофермент, катализирующий важнейшие окислительно-восстановительные реакции энергетического и пластического обмена. Заключенная в соединении НАДФ·Н химическая энергия является важным условием фотосинтеза, так как используется на восстановление CO2 и образование углеводов. Для восстановления одной молекулы НАДФ + в процессе фотосинтеза необходимы два электрона и два протона (причем донором протонов является вода).
Итак, на световой стадии фотосинтеза создается взаимодействие ФС II и ФС I, схематически изображаемое в виде зигзагообразной фигуры, которую называют Z-схемой. Обе фотосистемы (ФС II и ФС I) размещаются в тилакоидной мембране и работают в тесном контакте, обеспечивая друг другу бесперебойное функционирование.
Механизм фосфорилирования АТФ, сопряженный с работой электон-транспортной цепи на мембранах (хлоропластов и митохондрий), был расшифрован в 1961 году английским биохимиком Питером Митчеллом, лауреатом Нобелевской премии (1978).
Циклическое фосфорилирование. Выше был показан процесс образования АТФ, сопряженный с линейным перемещением электронов от ФС II к ФС I. Этот процесс называют нециклическим фосфорилированием. В процессе восстановления реакционного центра ФС I за счет электронного транспорта от ФС II между фотосистемами может возникнуть еще один поток электронов: от ферредоксина (Фд) фотосистемы I к комплексу цитохромов (b6/f) фотосистемы II. При этом НАДФ + не восстанавливается, а освобождающаяся энергия используется для фосфорилирования АДФ и образования макроэргической фосфатной связи АТФ. В итоге создается замкнутый электронный поток, при котором электроны, возбужденные в ФС I, не переходят на НАДФ + , а возвращаются в электрон-транспортную цепь на один из ее участков – «выше по течению». При этом перенос электронов по цепи сопровождается поступлением («перекачкой») протонов через тилакоидную мембрану. Возникающий в связи с этим электрохимический градиент обеспечивает синтез АТФ. Этот процесс называют циклическим фосфорилированием. Синтезированная АТФ переходит в строму хлоропласта.
Таким образом, во время световой фазы фотосинтеза благодаря энергии солнечного света синтезируется АТФ, образуется восстановленный НАДФ·Н и как побочный продукт выделяется кислород. В этом заключается основная функция световой фазы фотосинтеза. Она реализуется только при участии света и с помощью пигментов, размещенных в тилакоидной мембране хлоропластов.
Высокоэнергетические (макроэргические) соединения (АТФ, НАДФ·Н), образующиеся в ходе световой фазы, далее используются в темновой фазе фотосинтеза при ассимиляции CO2 в углеводы.