После формирования цветка происходят три важнейших взаимосвязанных процесса, без которых дальнейшее образование семян и плодов было бы невозможно. Это цветение, опыление и оплодотворение.
Цветение. Цветение – это готовность цветка к восприятию пыльцы. Обычно цветение начинается с раскрывания бутонов. У разных видов растений цветение наступает в определенном возрасте и в разное время года.
Однолетние и двулетние растения цветут и плодоносят один раз в жизни: однолетние – в первый (и единственный) год жизни, двулетние – на второй год. Многолетние растения цветут и приносят плоды многократно. Причем у древесных форм цветение, как правило, наступает в более зрелом возрасте. Например, дубы, растущие в лесу, впервые цветут в 80– 100-летнем возрасте. Однако среди многолетников существуют виды, которые зацветают только один раз в жизни, например агавы, бамбуки, некоторые пальмы.
У некоторых растений цветки способны раскрываться и закрываться несколько раз, в зависимости от внешних факторов (погоды, времени суток).
Существуют растения, у которых отсутствует выраженный сезон цветения, они образуют цветки в течение всего года. Такие растения называют ремонтантными (от франц. remontant – способный к многократному цветению). У них одновременно могут распускаться цветки и развиваться плоды. Как правило, это растения тропических широт (кофейное дерево, цитрусовые и др.).
Обычно цветение продолжается до опыления, но иногда – и до оплодотворения яйцеклеток.
Опыление. Опыление у покрытосеменных растений – это перенос пыльцевых зерен на рыльце пестика. Вспомните, что у голосеменных пыльца попадает непосредственно на семязачаток, открыто лежащий на чешуе шишки.
Перенос пыльцы осуществляется различными способами. Если пыльца данного цветка попадает на рыльце пестика этого же цветка, такое опыление называют самоопылением. Перенос пыльцы на рыльце пестика другого цветка называют перекресгпным опылением.
Самоопыление. Такой тип опыления характерен для небольшого числа цветковых растений. Ученые считают, что самоопыление возникло вторично, когда какие-то обстоятельства начали препятствовать осуществлению перекрестного опыления. Биологически самоопыление менее выгодно, поскольку при этом не происходит обмен генетической информацией между различными особями вида. Возникают гомозиготные популяции, не способные быстро адаптироваться к изменяющимся условиям окружающей среды.
Различают постоянное, случайное и вынужденное самоопыление. Постоянное самоопыление возникает, если перенос пыльцы с одного растения на другое невозможен в силу каких-то объективных причин. Для таких растений использование собственных-пыльцевых зерен – это единственная возможность образовать семена.
Случайное самоопыление происходит в том случае, когда при перекрестном опылении на рыльце пестика наряду с пыльцой других цветков также попадает пыльца из своего же цветка.
Вынужденное самоопыление происходит у некоторых покрытосеменных растений, для которых в обычных условиях характерно опыление чужой пыльцой. Если по каким-либо причинам перекрестного опыления не произошло, в последний момент перед окончанием цветения на рыльце попадает собственная пыльца.
Разновидностью самоопыления является клейстогамия (от греч. kleistos – замкнутый, gamos – брак), когда опыление собственной пыльцой происходит в нераспускающихся цветках.
Перекрестное опыление. Перекрестное опыление представляет собой перенос различными способами пыльцы из пыльника цветка одного растения на рыльце цветка другого растения. Этот тип опыления распространен у покрытосеменных растений гораздо шире, чем самоопыление. Биологически перекрестное опыление более благоприятно, чем самоопыление, потому что оно дает возможность объединять генетическую информацию разных особей, формировать новые сочетания аллелей, тем самым увеличивая генетическое разнообразие вида. Это способствует приспособлению вида к изменяющимся условиям обитания.
В процессе эволюции растения выработали ряд приспособлений, препятствующих самоопылению и способствующих перекрестному опылению. Рассмотрим некоторые из них.
У некоторых растений созревание пыльцы и подготовка рыльца к ее приему происходит не одновременно. Наиболее часто встречается ситуация, когда пыльники вскрываются и пыльца высыпается раньше, чем рыльце пестика того же цветка готово к ее восприятию. Так происходит у многих гвоздичных, колокольчиковых, сложноцветных и других растений.
Исключает возможность самоопыления двудомность, при которой пестичные и тычиночные цветки находятся на разных особях.
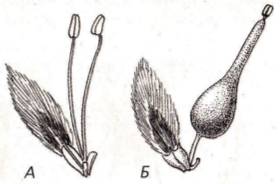
Существуют и другие приспособления, препятствующие самоопылению. У некоторых видов растений, например у примулы, на разных особях развиваются разные цветки: у одних цветков столбик пестика очень длинный, поэтому рыльце находится в зеве венчика, а тычинки с короткими тычиночными нитями в глубине; у других – наоборот. Такое явление называют гетеростилией (рис. 83). Понятно, что у таких растений возможно только перекрестное опыление.
Рис. 83. Гетеростилия у дербенника иволистного: А – длинностолбчатая форма; Б – короткостолбчатая форма
Перекрестное опыление может осуществляться различными способами. Условно их можно разделить на две группы: абиотическое опыление (при помощи ветра или воды) и биотическое (при помощи животных).
В роли опылителей могут выступать разные животные, поэтому выделяют энтомофилию (опыление насекомыми), орнитофилию (опыление птицами) и зоофилию (опыление млекопитающими).
Энтомофилия широко распространена в природе. Растения, опыляемые насекомыми, обладают разными приспособлениями, способствующими привлечению опылителей: яркая окраска цветка, обилие нектара, образование соцветий. Многих насекомых привлекает не только нектар, но и пыльца. Например, пчелы активно собирают пыльцу и переносят ее в гнездо на своих лапках. Там они складывают ее в ячейки сот и заливают медом. В таком виде пыльца может храниться очень долго. Пыльца у насекомоопыляемых растений, как правило, крупная, клейкая, с неровной поверхностью, что облегчает ее захват и перенос насекомыми.
Цветки привлекают насекомых не только внешним видом, но и запахом. Как правило, цветки, не имеющие яркой окраски, обладают сильным запахом. Важен запах и для привлечения ночных насекомых. Например, цветки душистого табака опыляются ночными бабочками. Поэтому днем они закрыты, а ночью, раскрываясь, распространяют вокруг сильный густой аромат. Цветки некоторых растений из семейства орхидных выделяют вещества, имитирующие запах самок насекомых в период размножения. Прилетевшие на запах самцы пытаются спариваться с цветком, который к тому же по цвету и форме тоже напоминает насекомое, а в результате покрываются пыльцой и переносят ее на другой цветок.
В процессе эволюции у насекомых и растений сформировалось огромное количество взаимных адаптаций. Эти адаптации иногда бывают настолько специальными, что некоторые виды покрытосеменных могут опыляться только определенными видами насекомых.
Орнитофилия, т. е. перенос пыльцы птицами, встречается гораздо реже. Птицы не воспринимают запах, поэтому наиболее привлекательны для них ярко и пестро окрашенные цветки, особенно красного цвета. Опылителями выступают колибри, нектарницы, медососы, попугаи лори и некоторые другие. В основном орнитофилия распространена в тропических и экваториальных областях.
Еще более редко встречается зоофилия. Летучие мыши по ночам и в сумерки опыляют крупные зеленовато-желтые или коричневые цветки с прочными околоцветниками и цветоножками, которые часто имеют неприятный запах, имитирующий выделения самих летучих мышей. Летучие мыши опыляют баобабы, мирты, агавы, бананы и некоторые другие растения, чаще всего деревья и кустарники, реже – травянистые растения. В опылении принимают участие и нелетающие животные: хоботноголовый кускус в Австралии, лемуры на Мадагаскаре, грызуны в Южной Америке.
Опыление ветром, или анемофилия, встречается у многих древесных и травянистых растений, но в целом ветроопыляемых растений гораздо меньше (около 10% от всего видового разнообразия). Цветки ветроопыляемых растений, как правило, имеют невзрачный околоцветник из пленок или чешуек, у некоторых видов цветки голые. Тычинки свешиваются из цветка наружу, их пыльники свободно раскачиваются ветром. Мелкая сухая легкая пыльца образуется в большом количестве. У ветроопыляемых растений цветки расположены значительно выше листьев (как, например, у злаков), или эти растения цветут до появления листьев (например, дуб).
При гидрофилии перенос пыльцы осуществляется с помощью воды. Например, у роголистника и зостеры цветки распускаются под водой. Их пыльцевые зерна тонкие, нитевидные, длиной до 2,5 мм. Наружная оболочка (экзина) практически редуцирована. Вероятность опыления в толще воды очень мала, поэтому такие растения размножаются в основном вегетативно.
Оплодотворение. Тем или иным способом пыльца попадает на рыльце пестика и удерживается на нем. С этого момента начинается новый этап – прорастание пыльцевого зерна (рис. 84). Плотная экзина прорывается, и из интины начинает формироваться пыльцевая трубка. Она проникает в рыльце и по столбику движется к завязи. В растущую пыльцевую трубку проникает ядро вегетативной клетки, образующей трубку, и два спермия. Достигнув завязи, пыльцевая трубка проникает внутрь семязачатка в зародышевый мешок через микропиле. Внутри зародышевого мешка оболочка пыльцевой трубки разрывается. Один спермий оплодотворяет яйцеклетку – возникает диплоидная зигота, из нее в дальнейшем развивается зародыш. Другой спермий сливается с диплоидным ядром центральной клетки, образуя клетку с тройным хромосомным набором (триплоидную), из которой затем формируется эндосперм – питательная ткань семени. Таким образом, у покрытосеменных растений в оплодотворении участвуют два спермия, т. е. осуществляется двойное оплодотворение. Механизм оплодотворения у цветковых растений бьп впервые открыт в 1898 г. русским ученым С. Г Навашиным.
Рис. 84. Жизненный цикл покрытосеменных растений