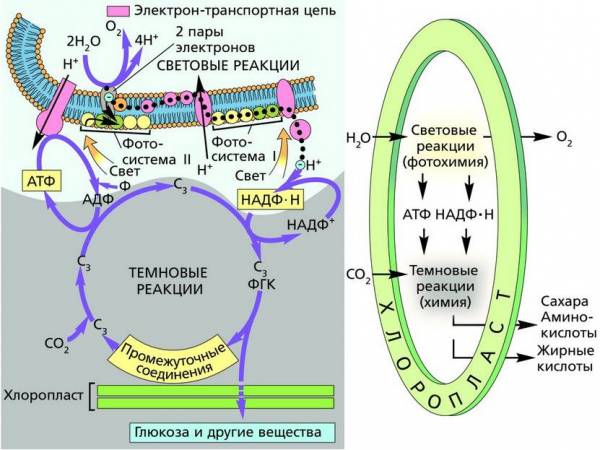
Клеточная мембрана
Биологические мембраны (биомембраны) – это сложные надмолекулярные структуры, ограничивающие содержимое клетки и большинство ее органоидов. Они представляют собой тончайшие, но достаточно прочные пленки толщиной 3,5–10 мкм, поэтому детально рассмотреть их структуру можно только с помощью электронного микроскопа. Биомембраны состоят преимущественно из белков и липидов, соотношение которых в разных мембранах может существенно различаться. Кроме того, в состав биомембран входят углеводы (приблизительно 10 %) и 20 % составляет прочно связанная вода.
Липиды (в основном это фосфолипиды, холестерин и гликолипиды) расположены в два слоя. Поэтому обычно говорят о бислойной структуре биомембран. В наибольшем количестве в мембранах присутствуют фосфолипиды. Это водонерастворимые органические молекулы, имеющие полярные «головки» (остатки фосфорной кислоты) и длинные неполярные «хвосты», представленные цепями жирных кислот. В бислое неполярные гидрофобные «хвосты» обращены друг к другу (погружены в толщу мембраны), а полярные гидрофильные «головки» ориентированы наружу. Биомембраны при ничтожно малой (по сравнению с протяженностью) толщине механически прочны, поскольку их матрикс образован двумя слоями сильно взаимодействующих гидрофобных структур.
Белки также играют важную роль в формировании структуры биомембраны. Они распределяются в ней неравномерно, например в мембранах эритроцитов имеется обширные участки липидного бислоя, практически свободные от белков.
Долгое время считалось, что биомембрана построена по модели «сэндвича», согласно которой она состоит из двух зеркально ориентированных липидных монослоев, а белки сплошным монослоем покрывают поверхность липидного бислоя.
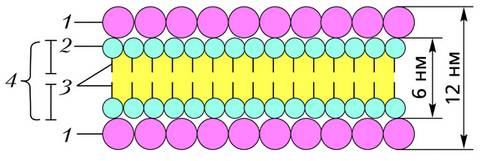 Схема молекулярного строения биологической мембраны (по К. Вилли, В. Детье): 1 – молекулы белка; 2 – гидрофильная часть молекул фосфолипидов; 3 – углеводородные цепи; 4 – двойной слой фосфолипидов
Схема молекулярного строения биологической мембраны (по К. Вилли, В. Детье): 1 – молекулы белка; 2 – гидрофильная часть молекул фосфолипидов; 3 – углеводородные цепи; 4 – двойной слой фосфолипидов
Однако в 70-е годы XX века большое внимание стало уделяться изучению динамических свойств мембран во взаимосвязи с выполняемыми ими функциями. Было обнаружено, что мембраны выполняют множество функций, которые модель «сэндвича» не была в состоянии объяснить.
В 1972 году американские ученые Г. Николсон и С. Сингер предложили новую – жидкостно-мозаичную модель строения биомембраны. Данные спектральных исследований, проведенных ими, указывали, что мембранные белки образуют глобулы, а не распределены в виде монослоя на поверхности липидного бислоя. Неполярные свойства мембранных белков наводили на мысль о наличии гидрофобных контактов между белками и внутренней неполярной областью липидного бислоя. В рамках предложенной Николсоном и Сингером модели биомембрана представляется как подвижный, текучий фосфолипидный бислой, в который погружены свободно диффундирующие белки. Белковые молекулы, пронизывающие в некоторых местах липидные слои мембраны, создают впечатление мозаики. Это отразилось в названии модели строения мембраны – жидкостно-мозаичная.
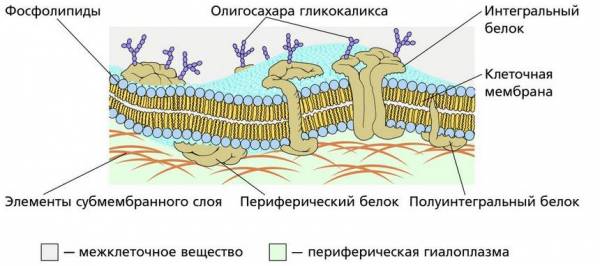 Строение поверхностного комплекса животной клетки
Строение поверхностного комплекса животной клетки
Мембрана, отделяющая клетку от внешней среды, называется клеточной или плазматической (цитоплазматической) мембраной. Клеточную мембрану растительных клеток обычно называют плазмалеммой. Строение клеточной мембраны у всех клеток одинаково. У прокариот (бактерий) это единственная мембрана в клетке. У эукариот мембраной отграничено не только содержимое клетки, но и клеточное ядро, а также различные органоиды: митохондрии, лизосомы, аппарат Гольджи, пероксисомы и др. Клеточные мембраны животных клеток богаты холестерином (до 30 %), а внутренние мембраны, отграничивающие части клетки – ядро и органонды – от цитоплазмы, содержат сравнительно много ненасыщенных жирных кислот (более жидких). Поэтому они оказываются менее жесткими.
Жидкостность мембраны определяется содержанием в фосфолипидах насыщенных и ненасыщенных жирных кислот и поведением самих молекул фосфолипидов, обладающих боковой подвижностью в пределах плоскости мембраны. Жесткость мембраны определяет холестерин.
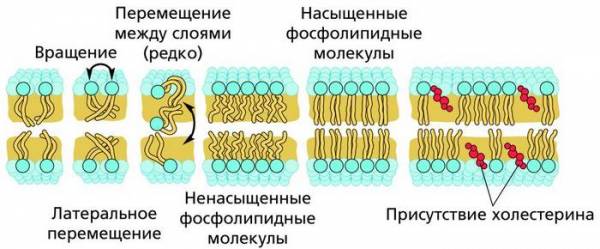 Подвижность фосфолипидного слоя в биомембране
Подвижность фосфолипидного слоя в биомембране
Вкрапление белковых молекул в мембрану происходит по-разному. Одни из них проходят через всю толщу мембраны так, что концы молекулы белка выходят наружу, – это интегральные, или трансмембранные, белки. Другие погружены в толщу фосфолипидных слоев мембраны лишь одним концом молекулы, а противоположный конец выходит наружу – их называют полуинтегральными белками. Многие погруженные белки мембран – ферменты. Третьи лежат снаружи мембраны, примыкая к ней, – это наружные, или периферические, белки. Некоторые белки могут находиться между фосфолипидными слоями.
Интегральные белки создают в мембране гидрофильные поры, через которые проходят водорастворимые вещества, поэтому их основной функций является транспорт веществ (среди них различают белки-переносчики и каналообразующие белки). Полуинтегральные белки выполняют преимущественно рецепторную функцию – воспринимают химические сигналы и передают их на определенные внутриклеточные (субмембранные) белки.
Четкая структурная организация и упорядоченность плазматической мембраны обусловливают ее жизненно важное свойство – полупронициаемость, то есть способность избирательно пропускать в клетку и выпускать из нее молекулы и ионы, в результате чего в клетке создается и поддерживаются определенная концентрация химических соединений.
Таким образом, клеточная мембрана – это органоид клетки, выполняющий важные функции: барьерную, транспортную, регуляторную и каталитическую. Она является основой поверхностного комплекса клетки.
Надмембранный слой
У эукариотических клеток плазматическая мембрана бывает покрыта еще дополнительными покровными структурами, образующими надмембранный слой и обладающими специфическими функциями. В животной клетке прилегающий непосредственно к плазматической мембране наружный слой называют гликокаликсом (от греч. glykys – сладкий и лат. callum – толстая кожа) . Он образован молекулами полисахаридов и олигосахаридов, связанных с мембранными белками или липидами. В гликокаликс входят также периферические белки и выступающие наружу рабочие части интегральных белков. Его толщина колеблется в пределах от 7 до 20 нм, причем она неодинакова даже в разных местах поверхностного слоя одной и той же клетки. Сложные углеводы гликокаликса представляют собой короткие линейные или разветвленные цепочки, в состав которых входят галактоза, манноза, фруктоза, сахароза, арабиноза и многие другие сахара. Выступая над мембраной, они, как и «веточки» молекул гликопротеинов гликокаликса, выполняют функцию специфических рецепторов и характеризуются большой индивидуальностью.
Другие структуры надмембранного слоя в животных клетках хорошо различаются как прилегающие к гликокаликсу периферические образования. Это могут быть слой слизи, хитина или еще одна мембраноподобная структура, которую называют перилеммой. У растений и грибов периферические структуры представлены клеточной стенкой. Клеточная стенка – это некий каркас, состоящий из многочисленных параллельно лежащих волокон, связанных между собой поперечными перемычками.
Клеточные стенки растений образованы преимущественно молекулами полисахарида целлюлозы, а у грибов – полисахарида хитина. Клеточные стенки растений пронизаны плазмодесмами (от греч. plasma – вылепленный и desmos – связь) – тонкими трубчатыми цитоплазматическими каналами, через которые соединяются между собой цитоплазмы соседних клеток. Диаметр этих каналов обычно составляет 20–40 нм. По оси канала из одной клетки в другую тянется цилиндрическая трубочка меньшего диаметра – десмотубула, просвет которой по данным электронной микроскопии сообщается с полостями эндоплазматической сети обеих смежных клеток.
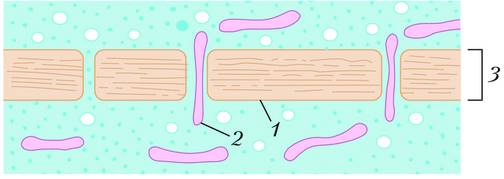 Схема строения клеточной стенки с плазмодесмами: 1 – плазматическая мембрана; 2 – мембрана десмотубулы; 3 – клеточная стенка
Схема строения клеточной стенки с плазмодесмами: 1 – плазматическая мембрана; 2 – мембрана десмотубулы; 3 – клеточная стенка
Линейные молекулы целлюлозы и хитина образуют высокоупорядоченные надмолекулярные структуры, вследствие чего клеточные стенки не растворяется в воде и очень устойчивы ко многим химическим веществам.
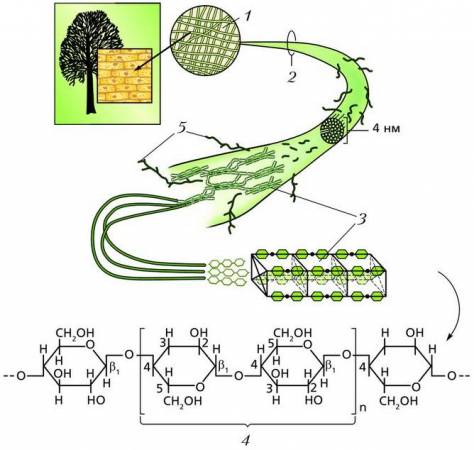 Строение клеточной стенки из линейных микрофибрилл целлюлозы: 1 – клеточная стенка; 2 – микрофибрилла целлюлозы; 3 – мономеры целлюлозы; 4 – дисахарид целлобиоза; 5 – молекулы гемицеллюлозы, выполняющие роль смазки в микрофибрилле
Строение клеточной стенки из линейных микрофибрилл целлюлозы: 1 – клеточная стенка; 2 – микрофибрилла целлюлозы; 3 – мономеры целлюлозы; 4 – дисахарид целлобиоза; 5 – молекулы гемицеллюлозы, выполняющие роль смазки в микрофибрилле
Белковые компоненты плазмалеммы и гликокаликса могут выполнять также ферментативную и транспортную функции.
Субмембранный слой
К поверхностному комплексу эукариотической клетки относят также ее субмембранный слой. В составе субмембранного слоя выделяют периферическую гиалоплазму и опорно-сократительную систему. Периферическая гиалоплазма является специализированной частью цитоплазмы, она расположена непосредственно под цитоплазматической мембраной. В ней располагается второй компонент субмембранного слоя – опорно-сократительная система, состоящая из разнообразных белковых нитевидных структур: тонких фибрилл (от лат. fibrilla – волоконце, ниточка), микрофибрилл, скелетных фибрилл и микротрубочек. Они связаны друг с другом белками и формируют опорно-сократительный аппарат клетки. Субмембранный слой взаимодействует с белками клеточной мембраны, которые, в свою очередь, связаны с надмембранным слоем. В результате информация, полученная в гликокаликсе или мембране, поступает к глубинным структурам клетки, в том числе к ядру, запуская тем самым сложные цепи биохимических реакций в ответ на поступившую информацию.
Таким образом, поверхностный комплекс клетки представляет собой многослойную, многофункциональную, структурно целостную систему. Это позволяет ему выполнять важные для клетки функции: изолирующую, транспортную, каталитическую, рецепторно-сигнальную и контактную.