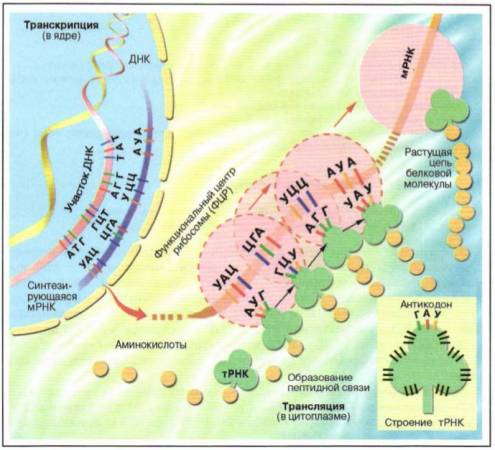
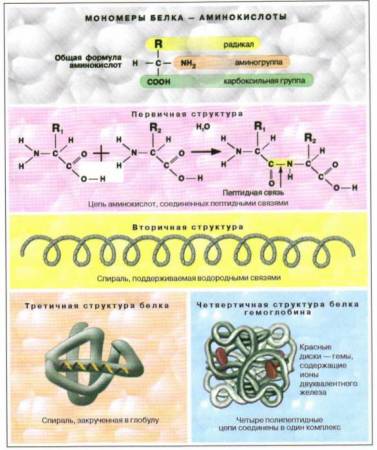
Почему закон независимого комбинирования признаков справедлив только для случаев, когда пары неаллельных генов находятся в разных парах хромосом?
Сцепление генов. Во всех примерах скрещивания, рассмотренных в предыдущих параграфах, имело место независимое комбинирование, поскольку интересующие нас пары генов находились в разных парах хромосом. Такие гены свободно сочетаются друг с другом в соответствии с закономерностью независимого комбинирования.
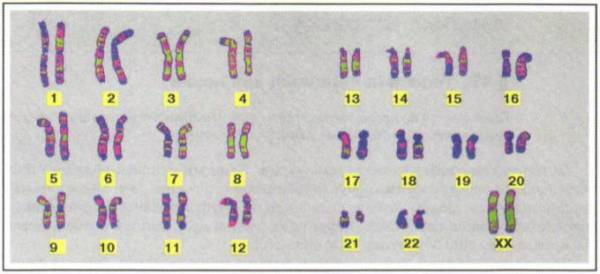
У различных организмов число генов может насчитывать десятки и сотни тысяч, число же хромосом весьма ограниченно, другими словами, число генов значительно превосходит число хромосом. Вот почему в каждой хромосоме располагается множество генов. Гены, локализованные в одной хромосоме, образуют группы сцепления; обычно они попадают в гамету вместе. Число групп сцепления равно числу пар хромосом, или гаплоидному числу хромосом (рис. 60). Это экспериментально подтверждено у всех хорошо изученных в этом отношении организмов.
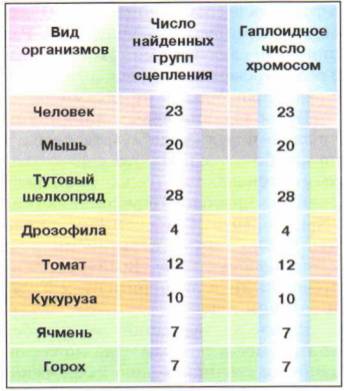 Рис. 60. Группы сцепления у разных видов организмов Сцепленное наследование. В случае расположения генов в одной хромосоме отмечается их сцепленное наследование – явление совместного наследования генов, локализованных в одной хромосоме (рис. 61).
Рис. 60. Группы сцепления у разных видов организмов Сцепленное наследование. В случае расположения генов в одной хромосоме отмечается их сцепленное наследование – явление совместного наследования генов, локализованных в одной хромосоме (рис. 61).
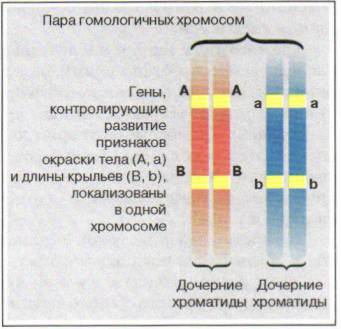 Рис. 61. Схема сцепления генов у дрозофилы
Рис. 61. Схема сцепления генов у дрозофилы
Сцепленное наследование изучал американский генетик Т. Морган на примере наследования признаков у плодовой мушки дрозофилы. (В XX в. дрозофила стала основным объектом генетических исследований.) Были проведены эксперименты по скрещиванию гомозиготных по доминантным аллелям мух (AABB) с серым телом и длинными крыльями и гомозиготных мух по рецессивным аллелям (aabb) с темным телом и редуцированными крыльями.
У гибридов F1 проявились только доминантные признаки – серая окраска тела и длинные крылья (согласно правилу единообразия гибридов первого поколения).
Затем гибриды F1 вновь скрестили с мухами, гомозиготными по рецессивным аллелям (aabb), т. е. провели анализирующее скрещивание.
Полученные результаты отличались от результатов дигибридного скрещивания. Ученые предположили, что пара неаллельных генов, т.е. генов, контролирующих развитие признаков окраски тела и длины крыльев, локализованы в одной хромосоме. В этом случае у дигетерозиготы возможно образование двух типов гамет (AB и ab) и появление двух типов потомков: с серым телом, длинными крыльями (AaBb) и темным телом с редуцированными крыльями (aabb). Причем таких гибридов должно быть поровну (по 50%), а формула расщепления – 1 : 1. Однако такого соотношения в эксперименте Моргана также не наблюдалось. Результаты анализирующего скрещивания были следующими: 41,5% серых длиннокрылых мух; 41,5% темных с редуцированными крыльями; 8,5% темных длиннокрылых; 8,5% серых с редуцированными крыльями.
Преобладание серых длиннокрылых и темных с редуцированными крыльями мух свидетельствовало о том, что гены A и B и a и b сцеплены, но не абсолютно. На это указывало появление мух с перекомбинированными признаками (серое тело, редуцированные крылья и темное тело, длинные крылья). Перекомбинация генов явилась следствием кроссинговера гомологичных хромосом у некоторых гамет (17 %) во время мейоза (см. § 18). Гаметы с перекомбинированными аллелями получили название кроссоверных гамет. Возникновение потомков с перекомбинироваиными признаками в экспериментах Т. Моргана – свидетельство нарушения сцепления генов.
В ходе исследований ученые убедились в том, что сцепление генов не всегда бывает полным. Оно может быть нарушено кроссинговером, приводящим к возникновению качественно новых (рекомбинантных) хромосом, вследствие чего образуются новые типы гамет. В итоге у потомков появляются новые комбинации признаков, отсутствующие у родительских особей.
Результаты экспериментов с мухой дрозофилой позволили сформулировать закон сцепленного наследования (или закон Моргана): гены, локализованные в одной хромосоме, часто наследуются совместно, образуя группу сцепления.
Хромосомная теория наследственности. Создание хромосомной теории наследственности стало одним из крупнейших теоретических обобщений генетики. Заслуга создания этой теории принадлежит Т. Моргану и сотрудникам его лаборатории К. Бриджесу, А. Стертеванту и Г. Мёллеру.
Согласно хромосомной теории наследственности, хромосомы являются носителями генов и представляют собой материальную основу наследственности. Каждая пара аллелей локализована в паре гомологичных хромосом, причем каждая из гомологичных хромосом несет по одному из пары аллелей.
Конкретные формы генов – аллели – занимают одно и то же место – локус – в гомологичных хромосомах.
Гены, расположенные в одной хромосоме, образуют группу сцепления и наследуются вместе. Число групп сцепления равно гаплоидному числу хромосом.
Нарушение сцепления – следствие кроссинговера в мейозе. Частота кроссинговера (процент кроссинговера) между двумя неаллельными генами, расположенными в одной хромосоме, пропорциональна расстоянию между ними: чем ближе расположены гены в хромосоме, тем теснее сцепление между ними и тем реже они разделяются при кроссинговере, и наоборот.
Закономерности, открытые научной школой Т. Моргана, были позднее подтверждены многочисленными исследованиями, проведенными на разных объектах.
Основные положения хромосомной теории наследственности заключаются в следующем.
- Гены расположены в хромосомах линейно.
- Каждый ген в хромосоме занимает определенное место (локус).
- Каждая хромосома представляет собой группу сцепления генов. Число групп сцепления у каждого вида организмов равно гаплоидному числу хромосом.
- Между гомологичными хромосомами может происходить обмен аллельными генами – кроссинговер.
- Расстояние между генами в хромосоме прямо пропорционально проценту кроссинговера. Это означает, что кроссинговер чаще может происходить между более удаленными друг от друга генами и реже– между близко расположенными генами.
Генетические карты. Принцип построения генетических карт (картирование) хромосом разработали Т. Морган и его ученики. В основу этого принципа положено представление о линейном расположении генов по всей длине хромосомы. Генетические карты хромосом представляют собой схему взаимного расположения генов, находящихся в одной группе сцепления. Иными словами, на генетических картах указан порядок расположения генов в хромосоме и относительные расстояния их друг от друга (рис. 62).
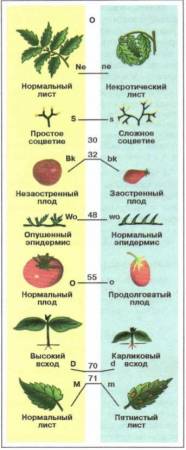 Рис. 62. Генетическая карта томата
Рис. 62. Генетическая карта томата
Генетические карты строится на основе прямого перевода частоты кроссинговера между генами в предполагаемые (условные) расстояния между ними в хромосоме. Единицей расстояния на генетической карте является морганида. Одна морганида соответствует 1% кроссинговера. Например, если частота кроссинговера между генами A и B равна 5%, то это означает, что они расположены в одной и той же хромосоме на расстоянии 5 морганид. Если же частота кроссинговера между генами A и C составляет 17%, то они разделены расстоянием в 17 морганид и т. д.
Ученым удалось картировать хромосомы многих организмов, в том числе человека. В настоящее время хромосомные и генетические карты широко используются на практике. Их применяют в селекционной практике и микробиологической промышленности, а также в медицинской генетике для диагностики ряда наследственных болезней и выявления мутаций. Без генетических карт невозможно развитие генной инженерии и молекулярной биологии.
Вопросы и задания
- Между какими хромосомами чаще происходит кроссинговер?
- Что называют группой сцепления? Чему равно число групп сцепления?
- Каковы положения хромосомной теории наследственности?
- Что такое генетическая карта? Какое значение имеет генетическое картирование?