СОДЕРЖАНИЕ
Связь ДНК с белками ядра клетки
ДНК в ядрах клеток эукариот обычно находится в тесном взаимодействии с ядерными белками разных групп: основными (гистоновыми) и кислыми (негистоновыми). Ассоциируясь, ДНК и белки образуют единый нуклеопротеидный комплекс –дезоксирибонуклеопротеид (ДНП). Процентное соотношение сухого веса всех указанных компонентов ДНП таково: ДНК – 35–40 %, гистоновые белки – 30–50 %, негистоновые белки – 4–33 %, то есть 40 % сухого веса составляет ДНК и около 60 % – белки.
Термин «хроматин» (от греч. chroma – цвет, краска) был введен в 1880 году немецким гистологом Вальтером Флеммингом (1843–1905). Хроматин легко окрашивается ядерными красителями при исследовании клеточного препарата с помощью светового микроскопа.
Хроматином обычно называют дисперсное (деспирализированное) состояние хромосом в интерфазе клеточного ядра эукариот (в неделящейся клетке). Но с начала деления ядра молекулы ДНК уже спирализированы (упакованы) в хромосомы. Одна молекула ДНК (точнее – комплекс ДНП) представляет собой одну хромосому. Спирализацию, или упаковку, ДНК осуществляют преимущественно гистоны.
Белки хроматина
В структурной организации ДНП центральную роль играют специфические белки – гистоны.
Гистоны – это относительно небольшие по молекулярной массе белки, присутствуют в ядрах клеток эукариот. Гистоновые белки богаты остатками аминокислот аргинина и лизина, определяющими их щелочные свойства. Практически все гистоны одинаковы, среди них насчитывают 5–7 типов молекул, обладающих сходными свойствами. Гистоны – это структурные белки, выполняющие важную роль – упаковку ДНК. Например, в растянутом состоянии двойная спираль ДНК, содержавшаяся в хромосоме человека, имеет длину в среднем 4 – 5 см, а будучи спирализованной в хромосоме при участии гистонов, измеряется долями микрометра. По сравнению с остальными белками, присутствующими в клетке, количество гистонов в клетке очень велико – оно почти равно массе ДНК, содержащейся в ядре, что свидетельствует об их активном участии в структурировании хроматина. Известно также, что гистоновые белки являются регуляторами биосинтеза нуклеиновых кислот (и ДНК, и РНК). Гистоны синтезируются в цитоплазме, но затем транспортируются в ядро и там связываются с ДНК во время ее репликации. При этом синтез гистонов и ДНК синхронизирован. Молекулярный комплекс ДНК-гистоны имеет форму особых субъединиц – нуклеосом (от лат. nucleus – ядро и греч. soma – тело).
Кроме того, в состав хроматина входит значительное количество других белков, объединяемых общим названием «негистоновые белки».
Негистоновые белки в сравнении с гистонами, наоборот, очень разнообразны. В хроматине насчитывается нескольких сотен типов их молекул. Среди них – ферменты, обеспечивающие процессы репликации ДНК, транскрипции, а также некоторые белки ядерного матрикса и матрикса ядрышка. Полагают, что негистоновые белки хроматина выполняют и некоторые регуляторные функции. Именно негистоновые белки участвуют в формировании самых высоких уровней упаковки ДНК.
Хроматин – это самый существенный, основной компонент ядерного аппарата клетки; из него образованы хромосомы клеток эукариот.
Формы упаковки ДНК
В процессе подготовки ядра клетки к делению, в интерфазе клеточного цикла, молекулы ДНК ассоциируются с белками и с их участием начинают «упаковываться», то есть скручиваться до минимальных размеров. Процесс упаковки хроматина (ДНП) до состояния размеров хромосомы называют процессом компактизации. Ведущая роль в организации расположения ДНК, ее компактизации и регулировании функциональных нагрузок принадлежит белкам.
Различают несколько структурных уровней компактизации хроматина в ядре клеток эукариот; от двухспиральной молекулы ДНК до ее суперупакованного состояния в хромосоме.
Как отмечалось выше, гистоны, синтезируемые в цитоплазме, транспортируются в ядро, где они связываются с длинной нитевидной цепью ДНК, начиная процесс ее упаковки. Среди ассоциированных с ДНК белков – гистоны HЗ и H4, содержащие большое количество аргинина, H2A и H2B, умеренно обогащенные лизином, и H1, представляющий собой не одну молекулу, а целый класс близкородственных белков, обогащенных лизином. Для всех классов гистонов, особенно для H1, характерно кластерное (групповое) распределение основных аминокислот на N- и C-концах молекулы. У гистона H1 обычно один N-конец связывается с другими гистоамии, а C-конец взаимодействует с ДНК. Эти гистоны и образуют нуклеосомы.
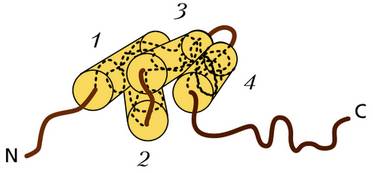 Схема третичной структуры белка гистона H1: спиральные участки (1–4) и фибриллярные C- и N-концы полипептидной цепи Нуклеосомы – структурные единицы хроматина, представляющие собой участки нити ДНК длиной около 200 пар оснований, уложенные на дисковидные гистоновые частицы диаметром около 10–11 нм. Они представляют собой октамер, или ядро, которое состоит из восьми молекул гистонов четырех типов (H2A, H2B, HЗ и H4, по две молекулы каждого). Вокруг гистонового октамера участок молекулы ДНК длиной в 140 пар нулкеотидов делает 1,75 витка. Диаметр сформированной таким образом нуклеосомы достигает 10 нм. Молекулы гистона (H1) не входят в структуру нуклеосом, но обеспечивают образование более высоких уровней упаковки ядерной ДНК.
Схема третичной структуры белка гистона H1: спиральные участки (1–4) и фибриллярные C- и N-концы полипептидной цепи Нуклеосомы – структурные единицы хроматина, представляющие собой участки нити ДНК длиной около 200 пар оснований, уложенные на дисковидные гистоновые частицы диаметром около 10–11 нм. Они представляют собой октамер, или ядро, которое состоит из восьми молекул гистонов четырех типов (H2A, H2B, HЗ и H4, по две молекулы каждого). Вокруг гистонового октамера участок молекулы ДНК длиной в 140 пар нулкеотидов делает 1,75 витка. Диаметр сформированной таким образом нуклеосомы достигает 10 нм. Молекулы гистона (H1) не входят в структуру нуклеосом, но обеспечивают образование более высоких уровней упаковки ядерной ДНК.
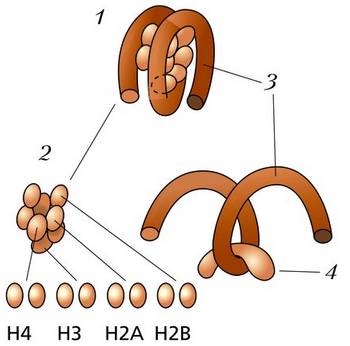 Схема строения нуклеосомы: 1 – нуклеосомная частица; 2– октамер, охватывающий четыре пары гистонов; 3 – фрагмент ДНК длиной в 146 пар оснований; 4 – гистон H1
Схема строения нуклеосомы: 1 – нуклеосомная частица; 2– октамер, охватывающий четыре пары гистонов; 3 – фрагмент ДНК длиной в 146 пар оснований; 4 – гистон H1
Формы упаковки (компактизации) хроматина (ДНП) в их последовательном усложнении показаны на схеме.
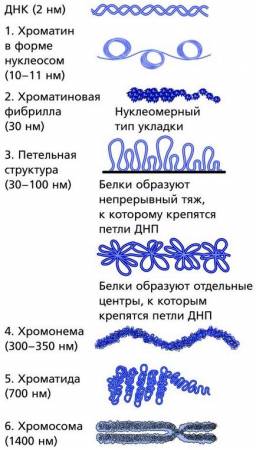 Уровни упаковки хроматина в ядре клеток эукариот (1-6 – уровни упаковки хроматина)
Уровни упаковки хроматина в ядре клеток эукариот (1-6 – уровни упаковки хроматина)
Уровни упаковки ДНК
Первым уровнем можно считать молекулярную форму ДНК в виде двойной спирали. Но поскольку она отдельно от белков в ядре практически не присутствует, чаще первым уровнем называют упаковку молекулы ДНК в нуклеосомной форме ДНП.
1. Первый уровень упаковки ДНП – нуклеосомная нить; она представляет собой структуру, напоминающую бусы на нитке, где в качестве бусин выступают нуклеосомы, а в качестве нитки – цепь ДНК. При этом толщина хроматиновой нити (ДНП) в нуклеосомах достигает 10–11 нм, что определяется фактически размерами самих нуклеосом.
Нуклеосомная нить («бусины на нитке»): 1 – ДНК; 2 – нуклеосома
2. Хроматиновая фибрилла – второй уровень упаковки хроматина. Представляет собой дальнейшую укладку нуклеосомной нити (бусин на нитке) в спираль с помощью гистона (H1). При формировании хроматиновой фибриллы происходит 40-кратная компактизация ДНП. Толщина такой фибриллы достигает уже 30 нм. Однако такого укорочения молекулы ДНК еще недостаточно даже для интерфазной хромосомы.
3. Петельная структура – третий уровень компактизации хроматина. Негистоновые белки образуют ось, или осевой скелет, – непрерывный тяж, к которому крепятся петли ДНП, имеющие форму хроматиновой фибриллы. На петельном уровне ДНК может достаточно легко освобождаться от упаковывающих ее белков, и на соответствующих участках становится возможной транскрипция (то есть синтез РНК).
4. Хромонема – форма хроматина четвертого уровня упаковки. Образуется путем конденсации (укладки) петельных фибрилл в отдельные участки – хромомерные (утолщенные) центры, которые у некоторых видов эукариот выглядят как узелки. При этом в самом конце интерфазы образуется серия динамических петель с большой толщиной (шириной). Толщина хромонемы уже достигает от 300 до 700 нм. В итоее достигается еще более плотная упаковка хроматина, прежде всего цепи ДНК.
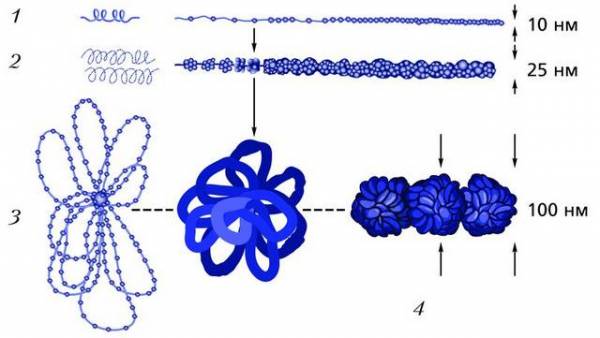 Схема начальных уровней компактизации хроматина: 1 – нуклеосомный; 2 – нуклеомерный; 3 – хромомерный (петлевой домен); 4 – хромонемный
Схема начальных уровней компактизации хроматина: 1 – нуклеосомный; 2 – нуклеомерный; 3 – хромомерный (петлевой домен); 4 – хромонемный
5. Перед началом деления ядра происходит удвоение хромонем, то есть их репликация и самовоспроизведение. При удвоении хромосомного аппарата обе сестринские хромонемы укладываются спирально или петлеобразно вместе, образуя хроматиду. В этом случае упакованная хромосомная нить достигает 700 нм в ширину.
6. Хромосома – шестая, последняя и самая суперспирализированная стадия упаковки. Состоящая из двух хроматид, она уплотнена, по сравнению с молекулой ДНК, в 100–500 раз. Ее толщина (ширина) достигает примерно 1400 нм. На стадии метафазы хромосомы уже видны в световой микроскоп.
Первые три уровня упаковки хроматина имеют место в интерфазном ядре и обозначаются на микрофотографиях как эухроматин, но с отдельными участками гетерохроматина. Начиная с третьего (петельного) уровня упаковка хроматина стабилизируется белками и разблокировка цепей ДНК происходит только на период считывания с них информации, то есть при синтезе РНК и редупликации ДНК. Однако еще до начала клеточного деления, в конце интерфазы, хроматин снова полностью спирализируется до уровня хромонем для обеспечения образования дочерних двойных цепей как основы хроматид, из которых впоследствии формируются хромосомы дочерних клеток.
Таким образом, процесс образования хромосом – сложное структурно-морфологическое преобразование, в основе которого лежит процесс компактизации структурных единиц в системе «молекула ДНК → хроматин (ДНП) → нуклеосома → хромонема → хромосома».