СОДЕРЖАНИЕ
Строение вируса
Обычно вирусы нельзя увидеть в световой микроскоп, так как отдельные вирусные частицы – вирионы – более чем вдвое меньше длины световой волны. Форма вирионов различна. Они могут быть нитевидными, сферическими, пулевидными, палочковидными, многоугольными, кирпичеобразными, кубическими, некоторые имеют кубическую головку и отросток. Вирусы имеют очень простое строение. Каждый вирион состоит лишь из нуклеиновой кислоты и белков.
В вирионах вирусов присутствует обычно только один тип нуклеиновой кислоты – либо РНК, либо ДНК. Причем и та и другая могут быть одноцепочечными и двухцепочечными, а ДНК – линейной или кольцевой. РНК в вирусах всегда только линейная, но может присутствовать и набор различных молекул РНК, каждая из которых несет определенную часть генетической информации, необходимой для репродукции. Особо следует подчеркнуть тот факт, что в царстве вирусов функцию носителя генетической информации выполняет не только ДНК, но и РНК.
В зависимости от типа нуклеиновой кислоты вирусы называют ДНК-содержащими и РНК-содержащими. На этой основе различают семейства вирусов. Они представлены на рисунке.
Каждый вирион состоит всего из двух частей – сердцевины, в которой находится ДНК или РНК, и окружающей белковой оболочки – капсида (от лат. capsa – вместилище, ящик, футляр). Белки защищают нуклеиновую кислоту, а также обеспечивают ферментативные процессы, обусловливающие изменения белкового состава капсида. Капсид состоит из определенным образом уложенных однотипных белковых молекул – капсомеров (или белковых субъединиц).
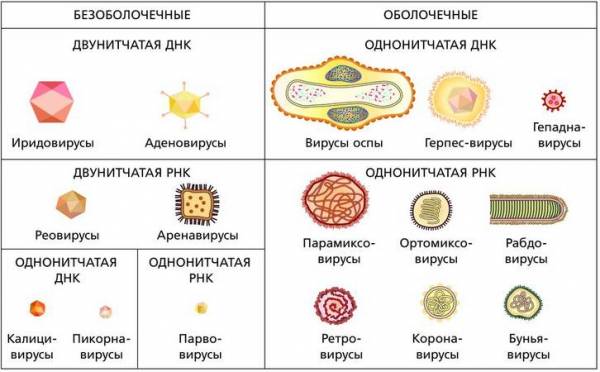 Семейства вирусов
Семейства вирусов
Обычно у вирусов наблюдается или спиральный тип укладки, или тип симметричного многогранника.
Модель вириона аденовируса: 1 – фибры; 2 – белки капсида; 3 – сердцевина (диаметр – 70-90 нм)
Все вирусы условно разделяют на простые и сложные. Простые вирусы состоят только из сердцевины с нуклеиновой кислотой и капсида. Сложные вирусы на поверхности белкового капсида имеют еще внешнюю оболочку (или суперкапсид), представленную двухслойной липопротеидной мембраной, углеводами и белками (ферментами). Эта внешняя оболочка обычно бывает построена из мембраны клетки-хозяина.
На поверхности капсида находятся различные выросты – шипы, или «гвоздики» (их называют фибрами), и отростки. Ими вирион прикрепляется к поверхности клетки, в которую он затем проникает. Следует отметить, что на поверхности вируса имеются еще специальные прикрепительные белки, связывающие вирион со специфическими группами молекул – рецепторами (от лат. recipio – получаю, принимаю), находящимися на поверхности клетки, в которую внедряется вирус. Одни вирусы прикрепляются к белковым рецепторам, другие – к липидам, третьи «узнают» углеводные цепочки среди белков и липидов. Вирусы «научились» узнавать чувствительные к ним клетки по наличию специальных рецепторов на клеточной поверхности «хозяев» в процессе эволюции.
Проникновение вируса в клетку
В 1979 году был описан универсальный механизм проникновения в эукариотическую клетку многих необходимых ей питательных веществ. Его назвали рецепторным эндоцитозом (от греч. endon – внутри и kytos – клетка). Эндоцитоз – это активное захватывание и поглощение клеткой инородных микроскопических объектов. Эндоцитоз сходен с пиноцитозом. Рецепторным он был назван потому, что объекты, прежде чем попасть в клетку, вначале прикрепляются к особым группам молекул на ее поверхности – рецепторам. Таким же путем попадают в клетку эукариот и вирусы.
Рецепторный эндоцитоз – основной путь проникновения вируса в клетку-хозяина.
Процесс проникновения вириона в клетку-хозяина включает несколько этапов: 1) прикрепление вируса к клеточным рецепторам; 2) образование на поверхности клетки из ямки вакуоли (эндоцитоз); 3) выход вируса из вакуоли в цитоплазму в результате слияния вирусной и клеточной мембран. Этот процесс представлен на рисунке.
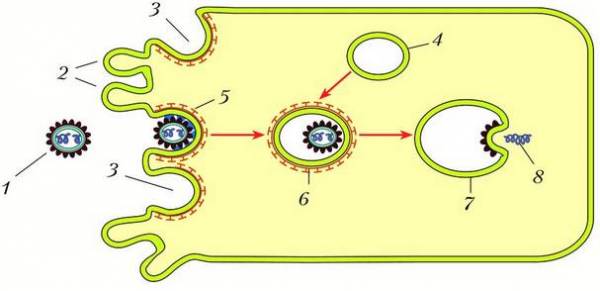 Проникновение вируса в клетку: 1 – вирусная частица; 2 – ворсинки на поверхности клетки; 3 – ямки с рецепторами; 4 – клеточная вакуоль; 5 – эндоцитоз; 6 – вакуоль, слившаяся с вирусом; 7 – слияние вирусной мембраны с мембраной вакуоли; 8 – выход генетического материала вируса из вакуоли
Проникновение вируса в клетку: 1 – вирусная частица; 2 – ворсинки на поверхности клетки; 3 – ямки с рецепторами; 4 – клеточная вакуоль; 5 – эндоцитоз; 6 – вакуоль, слившаяся с вирусом; 7 – слияние вирусной мембраны с мембраной вакуоли; 8 – выход генетического материала вируса из вакуоли
У прокариот существуют другие способы проникновения, например бактериофаг или фаг проникает в клетку бактерии без эндоцитоза. Имеющийся у бактериофага хвостовой отросток прикрепляется к клетке свободным концом. На конце отростка присутствует особый фермент – лизоцим. Как только фаг окажется на клетке, ее клеточная стенка под влиянием лизоцима растворяется и содержащаяся в головке фага ДНК перемещается в бактерию. Оболочка головки и отросток фага остаются снаружи клетки, ДНК сердцевины вируса – внутри. Этому процессу способствует и другой фермент – АТФаза. В клетке воспроизводятся новые особи фагов. Спустя некоторое время давление в клетке бактерии повышается от присутствия множества вирионов и под действием лизоцима клеточная стенка разрушается, фаги выходят в окружающую среду, заражая новые клетки бактерий.
В одних случаях выход вирусов происходит из уже погибшей клетки (опустошенной, разобранной паразитом на строительство новых, дочерних вирионов). В других случаях вирусы выходят из еще живой клетки вместе с частью ее цитоплазмы путем почкования через цитоплазматическую мембрану и клеточную стенку, одновременно приобретая суперкапсид. Выйдя из клетки, вирион способен снова проникать в другие клетки или пребывать в покоящемся состоянии. Существование вируса в двух формах – внеклеточной (покоящейся) и внутриклеточной (размножающейся) – рассматривают как две стадии цикла его индивидуального развития.
Выживание вирусов
Одним из удивительных свойств вирусов является их способность формировать кристаллические агрегаты как внутри пораженной клетки, так и вне ее. Важнейшими предпосылками для этого служат, во-первых, правильная кристаллическая форма вирусных частиц (вирионов); во-вторых, исключительная однородность вирионов одного вида по форме, величине и молекулярному весу.
Вирусные кристаллы образуются из десятков тысяч и даже миллионов вирусных частиц, причем кристаллизоваться может только идеально чистый препарат вируса. Ученые полагают, что кристаллическая структура скоплений (колоний) вирусов каким-то образом связана с функцией их выживания и размножения. Кристаллизация позволяет разместить в определенном объеме пространства максимальное количество вирионов (экономия пространства), и это, по-видимому, обеспечивает выживание частиц в экстремальных условиях. Об этом свидетельствуют многочисленные факты исключительной долголетней устойчивости кристаллических препаратов вирусов. Неясно лишь то, как способствует кристаллическая структура колоний размножению вирусов и какова энергетическая природа кристаллизации группировок вирионов.
Пребывание вирусов в кристаллическом состоянии некоторое время являлось одной из причин отнесения их к неживой природе.
Происхождение вирусов
Пути и механизмы возникновения вирусов пока еще не установлены. Существует несколько различных гипотез о происхождении вирусов. По одной из них предполагается, что вирусы являются наиболее ранними, древними доклеточными формами жизни, сохранившимися поныне благодаря паразитическому существованию в более прогрессивных клетках. Но эта гипотеза (1) не находит должного подтверждения. С одной стороны, для репродукции вирусов нужна клетка с ее метаболической системой, и, следовательно, это уже опровергает гипотезу доклеточного происхождения вирусов. С другой стороны, нуклеиновые кислоты и белки вирусов по уровню организации не уступают присутствующим в клеточных организмах. Генетический код вирусов аналогичен генетическому коду в ядрах клеток. По другой гипотезе (2) вирусы – это потомки каких-то доклеточных форм жизни, и на них природа «отрабатывала» пути эволюции в период зарождения жизни на Земле. В результате такой «отработки» возникли и вирусы, и прокариоты как особые ветви жизни.
Наиболее вероятной представляется гипотеза (3), согласно которой вирусы – это результат деградации таких клеточных организмов, как бактерии или клетки каких-то древних эукариот. Предполагают, что длительная эволюция этих клеточных организмов могла идти по пути формирования постоянно встречающегося паразитизма. Этот процесс сопровождался утратой белоксинтезирующего аппарата, автономной АТФ, цитоплазматической мембраны и цитоплазмы. При этом эволюция генетического материала, вероятно, шла по таким двум направлениям, как: 1) уменьшение объема информации и 2) упрощение процесса репродукции. При упрощении произошло вначале исключение этапа транскрипции списывания информации с ДНК на РНК), в результате чего РНК стала использоваться в качестве генетического материала.
Существует также гипотеза (4), по которой вирусы развились из отдельных органоидов эукариотической клетки – митохондрий и хлоропластов прокариотической клетки.
Есть и еще гипотеза (5), по которой вирусы появились после возникновения клетки и вместе с ней проделали длинный путь эволюции. Согласно этой гипотезе вирусы произошли от фрагментов клеточных нуклеиновых кислот – плазмид. Причем ДНК-содержащие вирусы приобрели свой генетический материал из клеточных ДНК, тогда как РНК-содержащие вирусы – из клеточных РНК. Полагают, что эти фрагменты клеточных нуклеиновых кислот вышли из-под контроля клетки, «оделись» собственной белковой оболочкой и приобрели способность заражать клетки, став патогенными агентами. Эта гипотеза носит название «взбесившихся генов», то есть клеточных генов, которые избавились от клеточного управления и стали паразитами самой же клетки.
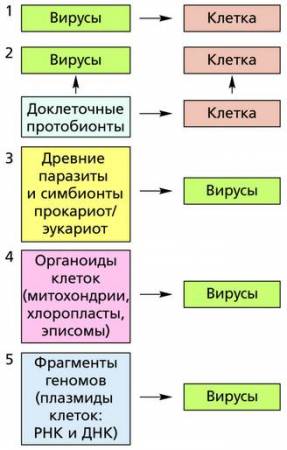 Схемы, иллюстрирующие гипотезы происхождения вирусов
Схемы, иллюстрирующие гипотезы происхождения вирусов
Как мы видим, ряд гипотез содержат предположения о том, что вирусы являются фрагментами живых клеточных организмов, утративших в процессе эволюции способность к самостоятельной репродукции, но сохранивших их некоторые наиболее существенные признаки, такие как наследственность, изменчивость, способность к адаптации и эволюции.