Темновая фаза фотосинтеза связана с реакциями фиксации углерода, которые проходят в строме хлоропласта и продолжаются в цитоплазме без непосредственного поглощения света. В процессе световой фазы фотосинтеза накапливается достаточно высокий уровень АТФ и НАДФ·Н. Однако сами по себе эти макроэргические соединения не способны синтезировать углеводы из CO2. Становится очевидным, что и темновая фаза фотосинтеза – сложный процесс, включающий большое количество последовательно идущих реакций, возможных только после осуществления световой фазы.
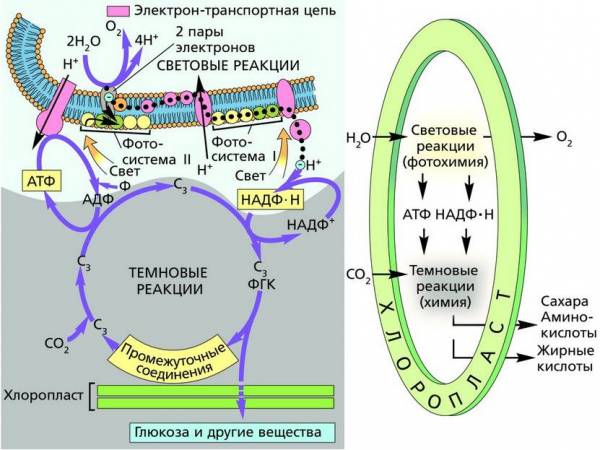 Общая схема взаимодействия световой и темновой фаз фотосинтеза
Общая схема взаимодействия световой и темновой фаз фотосинтеза
Существует несколько разных путей связывания CO2 в углеводы, встречающихся у растений разных экологических и систематических групп, но основным, характерным для всех растений, является так называемый C3-путь фотосинтеза, или цикл Кальвина.
Способ ассимиляции СО2 в углеводы, присущий всем растениям, был расшифрован только в середине XX века американским биохимиком Мэлвином Кальвином и его коллегами на примере одноклеточных зеленых водорослей (хлореллы и др.) и зеленых листьев шпината. Исследование этой проблемы продолжалось 10 лет – с 1946 по 1966 год. Вначале ученые вели поиск первичного акцептора CO2. После ряда экспериментов они установили, что первичную фиксацию CO2 осуществляет пятиуглеродный сахар – рибулозо-1,5-дифосфат РуДФ). Фиксация осуществляется следующим образом: сначала происходит присоединение CO2 к молекуле РуДФ. При этом образуется промежуточный продукт – очень неустойчивое шестиуглеродное соединение, из которого в присутствии воды образуются две молекулы трехуглеродного соединения – 3-фосфоглицериновой кислоты (3-ФГК). В этой реакции для связывания одной молекулы CO2 затрачивается три молекулы АТФ и две молекулы НАДФ·Н.
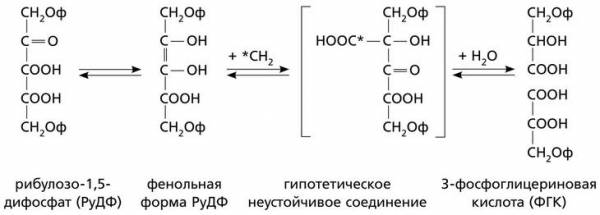 Схема первичной фиксации CO2
Схема первичной фиксации CO2
Реакцию фиксации углерода, открытую в 1948 году, катализирует очень крупный фермент из стромы хлоропласта – рибулозобисфосфаткарбоксилазаоксигеназа (сокращенно – РУБИСКО). Так как фермент РУБИСКО работает весьма медленно, необходимо, чтобы его молекул в хлоропластах было много. Действительно, этот фермент обычно составляет более 50 % общего количества белков хлоропластов. Многие исследователи утверждают, что это самый распространенный белок в живой природе.
Дальнейшие исследования лаборатории Кальвина способствовали установлению всех последующих реакций C3-пути фотосинтеза, обеспечивающих синтез углеводов.
За расшифровку механизма фиксации CO2 в процессе фотосинтеза М. Кальвин в 1961 году стал лауреатом Нобелевской премии по химии.
Цикл Кальвина
Цикл Кальвина состоит из трех стадий: карбоксилирования, восстановления и превращения.
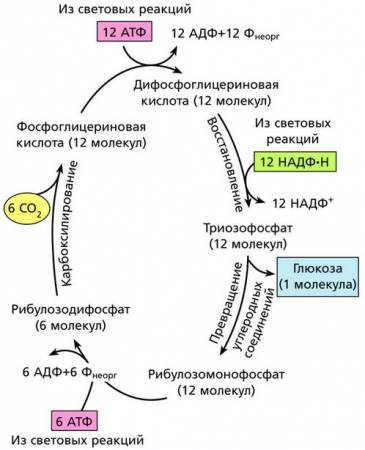 Упрощенная схема цикла Кальвина – пути фиксации углерода при фотосинтезе
Упрощенная схема цикла Кальвина – пути фиксации углерода при фотосинтезе
На первой стадии (карбоксилирование) фиксация углерода идет с участием ферментов и АТФ, полученной на световой фазе фотосинтеза; при этом образуются молекулы 3-фосфоглицериновой кислоты (3-ФГК). На второй стадии (восстановление) помимо АТФ используется НАДФ·Н. Здесь 3-ФГК восстанавливается до 3-фосфоглицеринового альдегида (З-ФГА), часть молекул которого идет на синтез 6-углеродного моносахарида (глюкозы или фруктозы). На третьей стадии (превращение) при повторении цикла другая часть молекул 3-ФГА используется для синтеза шестиатомного фосфорилированного моносахарида – фруктозо-1,5-дифосфата. Трехуглеродные фосфосахара вместе с множеством других продуктов метаболизма хлоропластов транспортируются в цитоплазму клетки, где образуют ди- и полисахариды (сахара, крахмал, целлюлозу или другие соединения).
В процессах темновой фазы фотосинтеза образуются углеводы – первичные органические вещества.
На определенном этапе темповой фазы фотосинтеза судьба трехуглеродных молекул 3-фосфоглицериновой кислоты может оказаться различной. Одни из них соединяются друг с другом и образуют шестиуглеродные сахара, которые, в свою очередь, могут полимеризоваться в крахмал, целлюлозу и др. Некоторые могут использоваться для синтеза аминокислот, карбоновых кислот, спиртов и пр. Но целый ряд молекул ФГК вовлекается в длинный ряд реакций, приводящих к превращению трехуглеродных молекул в молекулы пятиуглеродного сахара (РуДФ), которые могут снова ассимилировать углекислый газ и многократно повторять этот цикл до тех пор, пока растение живет и получает световую энергию. Все процессы темновой фазы фотосинтеза идут без непосредственного потребления света, но в них большую роль играют макроэргические соединения (АТФ и НАДФ·Н), образующиеся во время световой фазы фотосинтеза. Доказано, что для синтеза одной молекулы глюкозы в цикле Кальвина необходимы 12 молекул НАДФ·Н и 18 молекул АТФ, которые поставляются с тилакоидных мембран в результате фотохимических реакций световой фазы фотосинтеза.
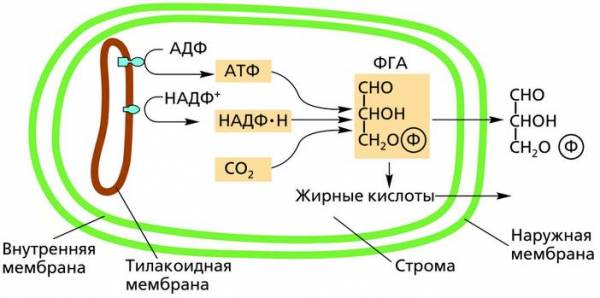 Схема синтеза 3-фосфоглицеринового альдегида (ФГА) в хлоропласте
Схема синтеза 3-фосфоглицеринового альдегида (ФГА) в хлоропласте
В процессе темновой фазы энергия макроэргических связей АТФ преобразуется в химическую энергию органических соединений – молекул углеводов. Это означает, что энергия солнечного света как бы консервируется в химических связях между атомами органических веществ, что имеет огромное значение для энергетики биосферы и жизнедеятельности всего населения нашей планеты.
Другие пути темновой фазы фотосинтеза
В настоящее время известны и другие пути ассимиляции углекислого газа наряду с системой его фиксации в цикле Кальвина (C3-пути фотосинтеза). Существует так называемый C4-путь ассимиляции углерода в фотосинтезе. Он может протекать при низких концентрациях CO2. Этот тип фиксации углекислого газа в фотосинтезе выработался в процессе эволюции у растений жарких, засушливых мест и наблюдается у кукурузы, сахарного тростника, проса, сорго, амаранта, лебеды, баклажанов и др., а также у растений, устойчивых к засолению почвы.
Существует и особый тип фотосинтеза у таких растений, как кактусы, молочаи, крассулы, каланхое, седумы и другие суккуленты, произрастающие в засушливых, безводных условиях. Эти растения запасают CO2 в виде органических кислот ночью, так как он поступает в клетки только тогда, когда открыты их устьица (днем они закрыты для предотвращения потери воды). Различные способы фиксации CO2 встречаются не только у растений. Особенности фотосинтеза у бактерий будут рассмотрены в следующем параграфе.
Факторы, влияющие на фотосинтез
На скорость фотосинтеза оказывают влияние многие факторы: количество (яркость) и продолжительность падающего света, его качественный состав; наличие влаги и минеральных веществ, поступающих в клетки; температура; концентрация CO2 и O2; суточные и сезонные ритмы; ветер и др.
У каждого вида растений при осуществлении ими фотосинтеза выработалась своя специфическая реакция на факторы внешней среды. При оптимальных условиях фотосинтез идет наиболее интенсивно. С ухудшением условий его темп обычно снижается. Например, для большинства высших растений умеренного пояса максимальная интенсивность фотосинтеза наблюдается при температуре 20–25 ºC. Однако, если концентрация CO2 в воздухе будет выше, температурный оптимум сместится до 35–38 ºC. Именно при этих температурах хорошо идут многие ферментативные реакции. Однако дальнейшее повышение температуры до 40–45 ºC приводит к резкому снижению скорости фотосинтеза. В то же время некоторые растения жарких пустынь способны осуществлять фотосинтез при температуре 58 ºC, а водоросли в слоевище лишайников в Антарктиде достигают максимальной интенсивности фотосинтеза при +5 ºC.
Оптимальные температурные условия для процесса фотосинтеза определяются прежде всего генотипом и условиями обитания растения. В летний день, в полуденные часы температура листьев может быть намного выше температуры воздуха. В таких условиях, как отмечает физиолог растений Сергей Семенович Медведев, у растений может наблюдаться «полуденная депрессия фотосинтеза» их клетки переполняются крахмалом, нарушается отток ассимилятов. От повреждений, наносимых солнечной радиацией, у растений имеется три уровня защиты. Первый – рассеивание энергии света в виде тепла. На втором уровне включается молекулярная система утилизации (каротиноиды, аскорбиновая кислота и др.) образующихся токсических продуктов. Если и второй уровень защиты не справляется, то токсические продукты вызывают повреждение в первую очередь тех молекул, которые особенно чувствительны к свету, например белка D1, входящего в состав фотосистемы II и обеспечивающего функции переносчика электронов из реакционного центра.