Нуклеиновые кислоты
Нуклеиновые кислоты – природные высокомолекулярные соединения, осуществляющие хранение, передачу и реализацию наследственной информации, обеспечивающей преемственность жизни в ряду поколений. Нуклеиновые кислоты представляют собой фосфорсодержащие биополимеры, которые отличаются неисчерпаемым разнообразием своих молекул. Мономерами нуклеиновых кислот служат нуклеотиды – соединения, в состав которых входят сахар (пентоза), фосфатная группа и азотсодержащее основание, связанные между собой ковалентными фосфодиэфирными связями, соединяющими пятый атом углерода пентозы одного нуклеотида с третьим атомом углерода пентозы соседнего нуклеотида.
Нуклеиновые кислоты были открыты в 1868 году швейцарским биохимиком И.Ф. Мишером в клетках, богатых ядерным материалом (в лейкоцитах и сперматозоидах лосося), в связи с чем и получили свое название (от лат. nucleus – ядро). Только в 1944 году было обнаружено, что очищенная ДНК одного бактериального штамма способна передавать свои свойства другому штамму. Однако этому открытию в то время биологи не придали большого значения, так как до 50-х годов считалось, что носителями наследственной информации служат белки. Лишь в начале второй половины XX века была доказана роль нуклеиновых кислот в хранении и передаче наследственной информации путем матричного биосинтеза белков.
В природе существуют два вида нуклеиновых кислот: дезоксирибонуклеиновая кислота (ДНК) и рибонуклеиновая кислота (РНК). ДНК и РНК – это длинные линейные полимерные цепочки (тяжи), состоящие из множества мономеров – нуклеотидов, последовательно соединенных с помощью фосфодиэфирной связи. Эта связь образуется за счет гидроксильных групп, одна из которых принадлежит фосфорной кислоте, соединенной с пятым углеродным атомом пентозы одного нуклеотида, а другая – третьему углеродному атому пентозы другого нуклеотида. Цепи нуклеиновых кислот насчитывают от нескольких десятков до сотен миллионов нуклеотидов, поэтому молекулярная масса нуклеиновых кислот составляет 10 5 – 10 10 .
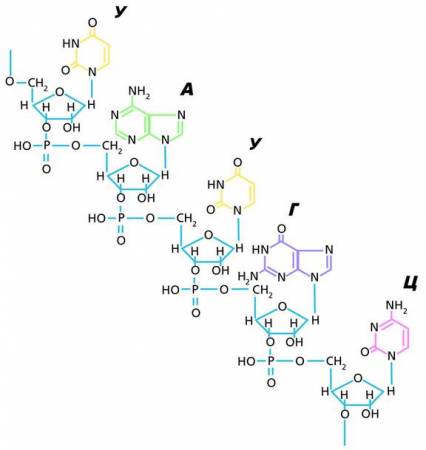 Фрагмент молекулы РНК
Фрагмент молекулы РНК
Молекула ДНК представлена двумя разнонаправленными полинуклеотидными цепями, закрученными одна вокруг другой в виде двойной спирали. Молекула РНК имеет одну полинуклеотидную цепь.
В 1953 году американский биохимик Джеймс Дьюи Уотсон и английский физик Фрэнсис Харри Крик, расшифровав структуру ДНК, построили ее модель. Ученые показали, что диаметр спирально закрученной двойной нити ДНК равен 2 нм, а на один виток– шаг спирали – приходится 10 пар нуклеотидов, что по протяженности занимает 3,4 нм. Все азотистые основания находятся внутри спирали, а пентозы и остатки фосфорной кислоты – снаружи. Пентозы с остатками фосфорной кислоты называют сахарофосфатным остовом. Двойная спираль достаточно прочно поддерживается посредством многочисленных водородных связей и гидрофобного взаимодействия.
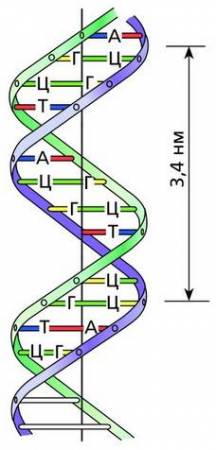 Модель молекулы ДНК (по Уотсону и Крику)
Модель молекулы ДНК (по Уотсону и Крику)
В 1962 году Дж. Уотсон, Ф. Крик и английский исследователь М. Уилкинс за раскрытие молекулярной структуры нуклеиновых кислот и ее значения в передаче генетической информации были удостоены Нобелевской премии.
С 1953 года роль ДНК как носителя наследственной информации, то есть как вещества генов, является основной биологической догмой в объяснении процессов жизни.
Нуклеотиды
Каждый нуклеотид содержит по одному остатку молекулы фосфорной кислоты и углевода (дезоксирибозу или рибозу), а также одно из четырех азотистых оснований: аденин, гуанин, цитозин и тимин (у ДНК), урацил (у РНК).
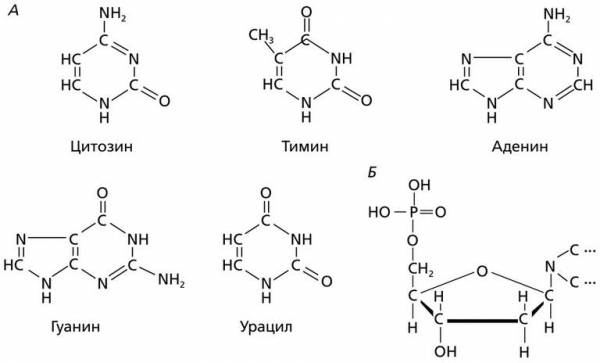 Химический состав нуклеотидов: А – структурные формулы азотистых оснований, входящих в состав нуклеотидов; Б – схема строения нуклеотида
Химический состав нуклеотидов: А – структурные формулы азотистых оснований, входящих в состав нуклеотидов; Б – схема строения нуклеотида
Азотистые основания, участвующие в образовании нуклеотидов и нуклеиновых кислот, относятся к группам пуриновых и пиримидиновых соединений. При этом пуриновые основания заметно крупнее пиримидиновых, поскольку состоят из двух гетероциклов. Тем не менее пуриновых оснований в ДНК ровно столько же, сколько и пиримидиновых. Это достигается тем, что в молекуле ДНК одно большое основание (А или Г) связывается с меньшим по размеру пиримидиновым основанием (Т или Ц), но входят они в ДНК в равных количествах. В одноцепочечной молекуле РНК пуриновых оснований обычно больше, чем пиримидиновых.
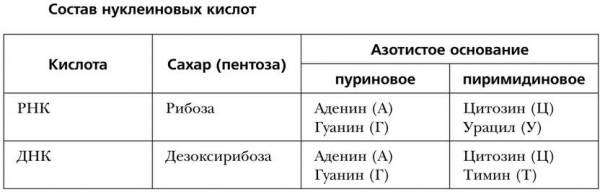 Состав нуклеиновых кислот
Состав нуклеиновых кислот
Название нуклеотида определяют по азотистому основанию и типу пентозы (например, гуаниловый дезоксирибонуклеотид, тимидиловый рибонуклеотид) .
Нуклеотиды выполняют много иных функций в клетке. Они запасают и переносят химическую энергию в составе молекул АТФ (аденин), ГТФ (гуанин), ЦТФ (цитозин); соединяются с другими биологически активными группами, образуя коферменты; участвуют во внутриклеточной передаче сигналов в качестве специфических сигнальных молекул. Однако их уникальное значение состоит именно в том, что они являются структурными компонентами молекул РНК и ДНК.
Структура ДНК
ДНК является наиболее высокомолекулярным органическим соединением из всех известных в природе. В клетках эукариот основная масса молекул ДНК находится в ядре в составе хромосом и имеет линейную форму. В прокариотических клетках единственная молекула ДНК имеет форму кольца. В виде небольших кольцевых молекул ДНК находится в некоторых органах клетки (митохондриях и пластидах). Будучи вытянутой в прямую линию, молекула ДНК может достигать 1–5 см.
Молекулы ДНК обычно построены по одному и тому же типу. Они состоят из двух полинуклеотидных цепей (нитей). Объединяются две цепи в единую полимерную молекулу ДНК водородными связями, возникающими между азотистыми основаниями нуклеотидов. При этом аденин (А) соединяется только с тимином (Т), а гуанин (Г) – с цитозином (Ц). Данные нуклеотиды в молекуле ДНК всегда располагаются парами: друг против друга.
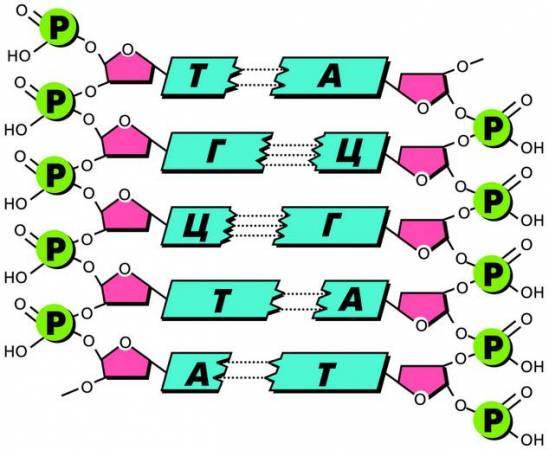 Основания ДНК, образующие комплементарные пары
Основания ДНК, образующие комплементарные пары
Способность к избирательному соединению нуклеотидов была названа правилом комплементарности или дополнительности, а соответствующие друг другу азотистые основания и нуклеотиды (А–Т и Г– Ц) – комплементарными.
Если известен порядок расположения нуклеотидов в одной цепи, то по принципу комплементарности сразу же становится ясным и порядок следования нуклеотидов в другой цепи. При этом следует учесть, что между А и Т замыкаются две водородные связи, а между Г и Ц – три, поэтому связь Г–Ц оказывается энергетически более прочной.
Антипараллельность молекул ДНК
Две цепи, участвующие в построении молекулы ДНК, всегда разнонаправлены, то есть имеют встречное расположение, так как З’-конец одной цепи соответствует 5′-концу другой цепи. Это обеспечивает антипараллельность цепей молекулы ДНК. Дело в том, что каждая пара нуклеотидов в каждой цепи связывается с последующей парой нуклеотидов посредством фосфорного остатка. При атом фосфорный остаток первой пары, связанный с третьим атомом углерода дезоксирибозы (3′-конец), соединяется с пятым атомом углерода дезоксирибозы второго нуклеотида (5′-конец). У второй цепи молекулы ДНК нуклеотид первой пары, наоборот, связан фосфорным остатком с пятым атомом углерода дезоксирибозы и третьим атомом углерода дезоксирибозы второго нуклеотида. Так же связываются все последующие пары нуклеотидов. В итоге у начала цепи всегда напротив нуклеотида с C3 дезоксирибозы (З’-конец) расположен нуклеотид с С5 дезоксирибозы (5′-конец).
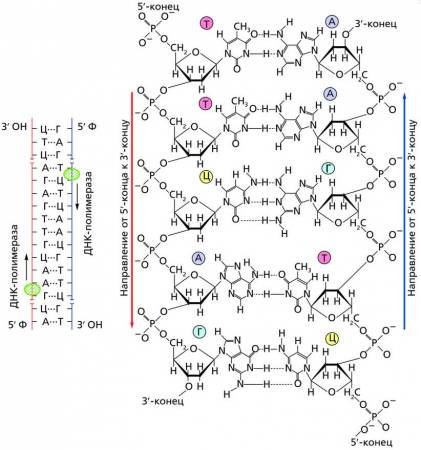 Антипараллельная ориентация цепочек ДНК
Антипараллельная ориентация цепочек ДНК
Антипараллельное расположение нитей ДНК имеет большое значение при ее репликации, так как ферменты (ДНК-полимеразы) могут передвигаться вдоль матричных цепей только от 5′-конца к 3′-концу.
Открытие двухцепочечной структуры молекулы ДНК имеет огромное значение. Это позволило установить основные принципы осуществления передачи наследственной информации. Поскольку последовательность нуклеотидов каждой цепи комплементарна последовательности нуклеотидов второй цепи, они несут одну и ту же генетическую информацию. При этом одна из двух цепей молекулы ДНК служит шаблоном, или матрицей, для образования новой цепи, а та, в свою очередь, тоже может служить матрицей для образования новой комплементарной цепи. Таким образом, генетическая информация может копироваться при разделении цепей.