Размножение составляет одну из важнейших характеристик сущности жизни. К размножению способны все без исключения живые организмы, от бактерий до млекопитающих. Существование каждого вида животных и растений, преемственность между родительскими особями и их потомством поддерживаются только благодаря размножению.
На молекулярном уровне процесс, который можно условно назвать размножением, выражается в уникальной способности ДНК к самоудвоению ее молекул. На уровне клетки к размножению путем деления способны такие органоиды, как митохондрии и хлоропласты. Клетки одноклеточных и многоклеточных организмов размножаются делением. Формы размножения организмов очень разнообразны и сложны, но в основе всех способов бесполого и полового размножения лежит деление клетки.
Митоз
Основной способ деления эукариотических клеток – митоз (греч. «митос» – нить).
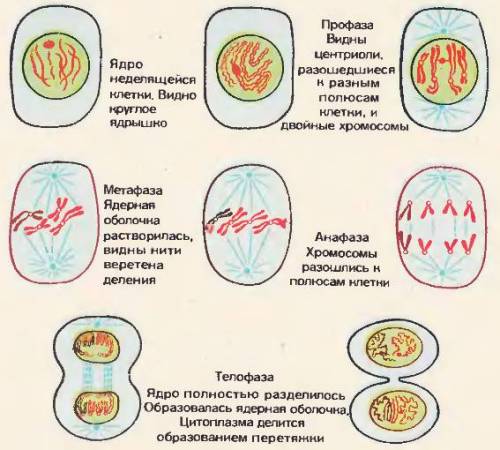 Митоз клетки животных
Митоз клетки животных
Подготовка клеток к делению начинается в период интерфазы. Один из важнейших подготовительных процессов – синтез ДНК, т. е. удвоение ее молекулы, происходящее в середине периода интерфазы. Таким образом. к митозу приступают клетки, содержащие удвоенное после синтеза количество ДНК.
В митозе различают четыре фазы: профазу, метафазу, анафазу, телофазу. Во время профазы в ядре появляются тонкие нити. Эти нити – хромосомы. В профазе они спирализуются и потому начинают укорачиваться и утолщаться.
К концу профазы все хромосомы становятся хорошо видны в световой микроскоп, можно определить их размеры, форму, строение, число. Каждая хромосома – это удлиненное плотное тельце, состоящее и нескольких частей, отделенных друг от друга перетяжками. Различают первичную перетяжку, или центромеру (греч. «мерос» – часть). На хромосоме может быть и вторичная перетяжка.
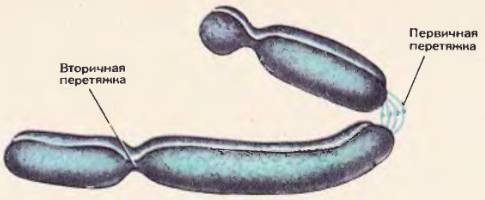 Строение хромосомы
Строение хромосомы
Каждая хромосома состоит из двух свернутых в спираль нитей (молекул) ДНК, которые называют хроматидами или дочерними хромосомами.
Во время профазы центриоли, а их две в каждой клетке, расходятся к противоположным полюсам клетки и между ними образуется веретено деления. В конце профазы ядерная оболочка растворяется и хромосомы свободно располагаются в цитоплазме, ядрышки исчезают.
За профазой следует метафаза митоза. Во время метафазы завершается образование веретена деления и хромосомы располагаются в экваториальной плоскости веретена. Хромосомы образуют метафазную пластинку, и каждая хромосома прикрепляется своим центральным участком (центромерой) к одной из нитей веретена. У каждой хромосомы происходит отделение, обособление хроматид друг от друга.
Когда все хромосомы оказываются прикрепленными к нитям веретена, хроматиды каждой хромосомы начинают расходиться к полюсам клетки: к одному полюсу отходит одна хроматида, к противоположному – другая. Начало расхождения хроматид к полюсам клетки означает, что наступила следующая фаза митоза – анафаза. Во время анафазы хроматиды (дочерние хромосомы) расходятся к полюсам клетки. Движение хромосом осуществляется благодаря нитям веретена, которые сокращаются и растягивают дочерние хромосомы от экватора к полюсам клетки. При движении хромосом используется энергия АТФ.
Последняя фаза митоза – телофаза. Во время телофазы приблизившиеся к полюсам клетки хромосомы начинают раскручиваться и снова приобретают форму длинных нитей, переплетающихся друг с другом, что характерно для неделящегося ядра [86]. В дочерних ядрах вновь образуется ядерная оболочка, формируется ядрышко и полностью восстанавливается характерное для интерфазы строение ядра. На протяжении телофазы происходит и деление цитоплазмы, в результате которого две дочерние клетки отделяются друг от друга. Эти клетки по строению полностью сходны с материнской.
Продолжительность митоза
В жизненном цикле клетки мигов занимает значительно более короткий промежуток времени, чем интерфаза.
В большинстве клеток весь процесс митоза, от профазы до завершения телофазы, занимает 1-2 ч, а интерфаза во много раз продолжительнее.
Биологическое значение митоза
В результате митоза каждая дочерняя клетка получает точно такие же хромосомы, какие имела материнская клетка. Число хромосом в обеих дочерних клетках равно числу хромосом материнской клетки. Следовательно, биологическое значение митоза заключается в строго равномерном распределении хромосом между ядрами двух дочерних клеток. Это значит, что мигов обеспечивает точную передачу наследственной информации каждому из дочерних ядер.
Если нарушается нормальный ход митоза и в дочерней клетке хромосом окажется меньше или больше, чем в материнской, то это приведет либо к гибели, либо к существенным изменениям в жизнедеятельности клетки – к возникновению мутаций.
Число хромосом
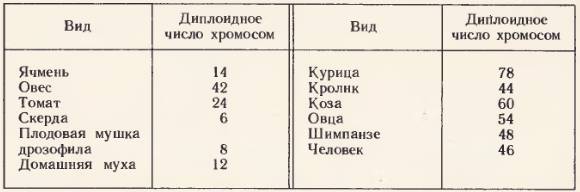
Основу генетического критерия вида составляет число хромосом в клетке, которое постоянно для каждого вида живых организмов.
Данные о количестве хромосом у некоторых видов растений и животных приведены в таблице.
Хромосомы, содержащиеся в ядре одной клетки, всегда парные, т. е. в ядре имеются две одинаковые, или гомологичные, хромосомы, которые составляют одну пару. Так, 46 хромосом человека образуют 23 пары, в каждой паре две одинаковые хромосомы. Хромосомы разных пар отличаются друг от друга размерами, формой, местами расположения первичных и вторичных перетяжек.
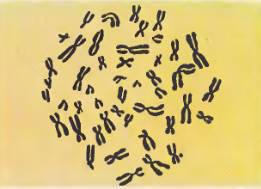 Диплоидный набор хромосом человека
Диплоидный набор хромосом человека
Совокупность хромосом, содержащихся в одном ядре, носит название хромосомного набора. Хромосомный набор характерен для каждого вида организмов. Примеры некоторых наборов хромосом приведены на рисунке.
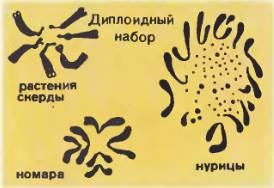 Диплоидный набор хромосом в клетках
Диплоидный набор хромосом в клетках
В любом многоклеточном организме различаются две категории клеток: соматические (неполовые), которые входят в состав всех тканей и органов тела, и половые клетки. Ядра соматических клеток содержат диплоидный (двойной) набор хромосом. Ядра половых клеток содержат гаплоидный, т. е. одинарный, набор хромосом. Так, если диплоидный набор ржи включает 14 хромосом, то гаплоидный набор имеет 7 хромосом. Если диплоидный набор человека содержит 46 хромосом, то гаплоидный набор равен 23 хромосомам. В гаплоидном наборе от каждой пары остается только одна хромосома.
Число хромосом и диплоидного и гаплоидного набора постоянно для каждого вида организмов.
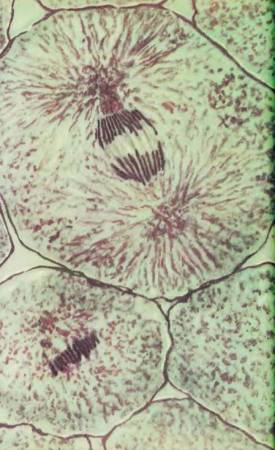 Микроскопическая фотография митоза клетки животных
Микроскопическая фотография митоза клетки животных
Профаза (внизу) — хромосомы свободно расположены в цитоплазме; метафаза (слева) — хромосомы упорядоченно лежат в одной плоскости; анафаза (вверху) — хромосомы расходятся к полюсам клетки.