Клетка любого организма представляет собой целостную живую систему. Она состоит из трех неразрывно связанных между собой частей: оболочки, цитоплазмы и ядра. Оболочка клетки осуществляет непосредственное взаимодействие с внешней средой и взаимодействие с соседними клетками (в многоклеточных организмах).
Оболочка клеток. Оболочка клеток имеет сложное строение. Она состоит из наружного слоя и расположенной под ним плазматической мембраны. Клетки животных и растений [49], [50], [51] различаются по строению их наружного слоя. У растений, а также у бактерий, сине-зеленых водорослей и грибов на поверхности клеток расположена плотная оболочка, или клеточная стенка. У большинства растений она состоит из клетчатки.
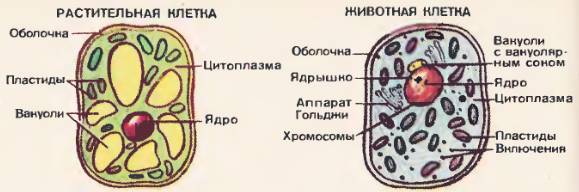 Рис. 49. Схемы строения клеток по данным светового микроскопа
Рис. 49. Схемы строения клеток по данным светового микроскопа 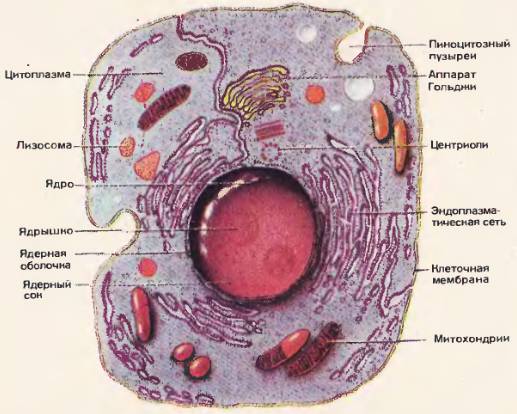 Рис. 50. Схема строения животной клетки по данным электронного микроскопа
Рис. 50. Схема строения животной клетки по данным электронного микроскопа 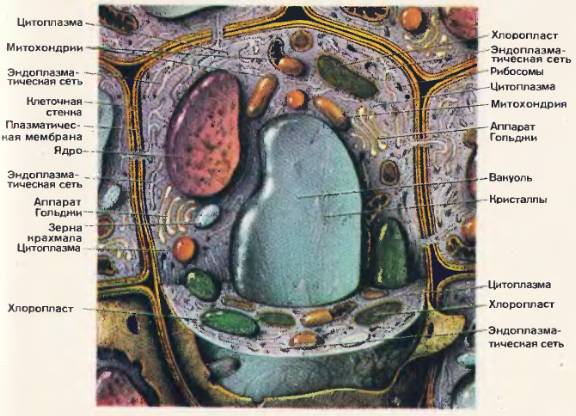 Рис. 51.
Рис. 51.
Клеточная стенка играет исключительно важную роль: она представляет собой внешний каркас, защитную оболочку, обеспечивает тургор растительных клеток; через клеточную стенку проходит вода, соли, молекулы многих органических веществ.
Наружный слой поверхности клеток животных [49], [50] в отличие от клеточных стенок растений очень тонкий, эластичный. Он не виден в световой микроскоп и состоит из разнообразных полисахаридов и белков. Поверхностный слой животных клеток получил название гликокаликс.
Гликокаликс выполняет прежде всего функцию непосредственной связи клеток животных с внешней средой, со всеми окружающими ее веществами. Имея незначительную толщину (меньше 1 мкм), наружный слой клетки животных не выполняет опорной роли, какая свойственна клеточным стенкам растений. Образование гликокаликса, так же как и клеточных стенок растений, происходит благодаря жизнедеятельности самих клеток.
Плазматическая мембрана. Под гликокаликсом и клеточной стенкой растений расположена плазматическая мембрана (лат. «мембрана» – кожица, пленка), граничащая непосредственно с цитоплазмой [52]. Толщина плазматической мембраны около 10 нм, изучение ее строения и функций возможно только с помощью электронного микроскопа.
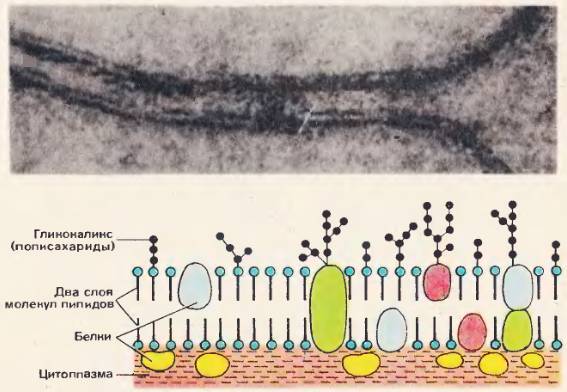 Рис. 52. Строение плазматической мембраны (электронно-микроскопическая фотография — вверху)
Рис. 52. Строение плазматической мембраны (электронно-микроскопическая фотография — вверху)
В состав плазматической мембраны входят белки и липиды. Они упорядоченно расположены и соединены друг с другом химическими взаимодействиями. По современным представлениям молекулы липидов в плазматической мембране расположены в два ряда и образуют сплошной слой [52]. Молекулы белков не образуют сплошного слоя, они располагаются в слое липидов, погружаясь в него на разную глубину, как это показано на рисунке 52.
Молекулы белка и липидов подвижны, что обеспечивает динамичность плазматической мембраны.
Плазматическая мембрана выполняет много важных функций, от которых зависит жизнедеятельность клеток. Одна из таких функций заключается в том, что она образует барьер, отграничивающий внутреннее содержимое клетки от внешней среды. Но между клетками и внешней средой постоянно происходит обмен. веществ. Из внешней среды в клетку поступает вода, разнообразные соли в форме отдельных ионов, неорганические и органические молекулы. Они проникают в клетку через очень тонкие каналы плазматической мембраны. Во внешнюю среду выводятся продукты, образованные в клетке. Транспорт веществ – одна из главных функций плазматической мембраны.
Через плазматическую мембрану из клетки выводятся продукты обмена, а также вещества, синтезированные в клетке. К числу их относятся разнообразные белки, углеводы, гормоны, которые вырабатываются в клетках различных желез и выводятся во внеклеточную среду в форме мелких капель.
Клетки, образующие у многоклеточных животных разнообразные ткани (эпителиальную, мышечную и др.), соединяются друг с другом плазматической мембраной. В местах соединения двух клеток мембрана каждой из них может образовывать складки или выросты, которые придают соединениям особую прочность [53].
 Рис. 53. Электронно-микроскопическая фотография мембран двух соседних клеток (Видны складки и выросты наружной мембраны, увеличивающие прочность соединения клеток. Увел. 30 000 раз.)
Рис. 53. Электронно-микроскопическая фотография мембран двух соседних клеток (Видны складки и выросты наружной мембраны, увеличивающие прочность соединения клеток. Увел. 30 000 раз.)
Соединение клеток растений обеспечивается путем образования тонких каналов, которые заполнены цитоплазмой и ограничены плазматической мембраной. По таким каналам, проходящим через клеточные оболочки, из одной клетки в другую поступают питательные вещества, ионы, углеводы и другие соединения.
На поверхности многих клеток животных, например различных эпителиев, находятся очень мелкие тонкие выросты цитоплазмы, покрытые плазматической мембраной, микроворсинки. Наибольшее количество микроворсинок находится на поверхности клеток кишечника, где происходит интенсивное переваривание и всасывание переваренной пищи.
Фагоцитоз. Крупные молекулы органических веществ, например белков и полисахаридов, частицы пищи, бактерии поступают в клетку путем фагоцитоза (греч. «фагео» – пожирать). В фагоцитозе непосредственное участие принимает плазматическая мембрана [54]. В том месте, где поверхность клетки соприкасается с частицей какого-либо плотного вещества, мембрана прогибается, образует углубление и окружает частицу, которая в «мембранной упаковке» погружается внутрь клетки. Образуется пищеварительная вакуоль и в ней перевариваются поступившие в клетку органические вещества.
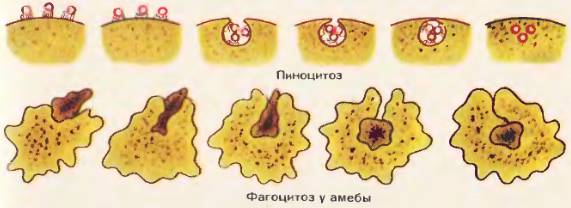 Рис. 54. Схема пиноцитоза. Фагоцитоз у амебы
Рис. 54. Схема пиноцитоза. Фагоцитоз у амебы
Фагоцитоз широко распространен в мире животных. Путем фагоцитоза питаются амебы, инфузории и многие другие простейшие. У позвоночных животных и человека к активному фагоцитозу способны только немногие клетки, например лейкоциты. Эти клетки поглощают бактерии, а также разнообразные твердые частички, случайно попавшие в организм, защищая его таким образом от болезнетворных микроорганизмов и посторонних частиц. Клеточная стенка растений, бактерий и сине-зеленых водорослей препятствует фагоцитозу и потому этот путь поступления веществ в клетку у них практически отсутствует.
Пиноцитоз. Через плазматическую мембрану в клетку проникают и капли жидкости, содержащие в растворенном и взвешенном состоянии разнообразные вещества.
Поглощение жидкости в виде мелких капель напоминает питье, и это явление было вызвано пиноцитозом (греч. «пино» – пью). Процесс поглощения жидкости сходен с фагоцитозом. Капля жидкости [54] погружается в цитоплазму в «мембранной упаковке». Органические вещества, попавшие в клетку вместе с водой, начинают перевариваться под влиянием ферментов, содержащихся в цитоплазме.
Пиноцитоз широко распространен в природе и осуществляется клетками животных, растений, грибов, бактерий и сине-зеленых водорослей.
Процессы фаго- и пиноцитоза, транспорт ионов и молекул происходит с затратой энергии, которая образуется в клетке.
1. Сравните строение оболочки растительных и животных клеток. 2. Как происходит поступление веществ а клетку? 3. Каковы основные функции гликокаликса, клеточкой стенки и плазматической мембраны?