Наследственная информация – это генетическая информация, которая передается по наследству от родителей потомству. Она обеспечивает преемственность поколений и непрерывность существования биологических видов.
У всех клеточных организмов и ряда вирусов единственную, но очень важную роль носителя наследственной информации играет ДНК.
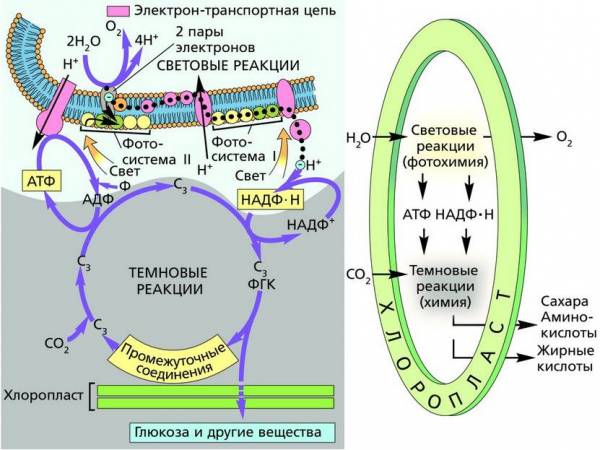
Считывание генетической информации и ее реализация в процессах биологического синтеза осуществляются посредством различных ферментных комплексов и разных молекул РНК. Однако главным компонентом– хранителем наследственной информации являются молекулы ДНК. Это определяется тем, что двухцепочечная молекула ДНК способна к самоудвоению, обеспечивая тем самым точное воспроизведение идентичных дочерних молекул. Кроме того, ДНК служит матрицей для синтеза всех видов РНК. Полагают, что на самых ранних этапах возникновения жизни эту функцию выполняла РНК, но позднее она перешла к ДНК.
Наследственная информация закодирована последовательностью нуклеотидов молекул ДНК (у некоторых вирусов – РНК). Она содержит сведения о строении всех ферментов (а их в природе существует около 10 тыс.), всех структурных белков и РНК клетки, а также о системах регуляции их синтеза.
Итак, именно ДНК определяет наследственность организмов, то есть воспроизводящийся в поколениях набор белков и связанных с ними признаков. Биосинтез белков является центральным процессом живой материи, а нуклеиновые кислоты обеспечивают его, с одной стороны, программой, определяющей весь набор и специфику синтезируемых белков, а с другой – механизмом точного воспроизведения этой программы в поколениях.
Понятие о репликации
Самоудвоение, или репликация (от лат. replicatio – повторение), – это процесс самовоспроизведения макромолекул нуклеиновых кислот, обеспечивающий точное копирование генетической информации. В основе этого механизма лежит ферментативный синтез ДНК на матрице ДНК или РНК на матрице РНК (у РНК-содержащих вирусов). Важное место среди ферментов, осуществляющих этот процесс, занимает ДНК-зависимая ДНК-полимераза, или просто ДНК-полимераза.
Напомним, что перед каждым делением клетки, в интерфазе клеточного цикла, происходит самоудвоение молекул ДНК. Процесс самоудвоения ДНК осуществляется с помощью фермента ДНК-полимеразы при участии четырех типов мономеров – дезоксирибонуклеозидтрифосфатов (дАТФ, дТТФ, дГТФ и дЦТФ), являющихся «поставщиками» и нуклеотидов для сборки новой молекулы ДНК, и необходимой для этого энергии, носителем которой служит трифосфатная группа.
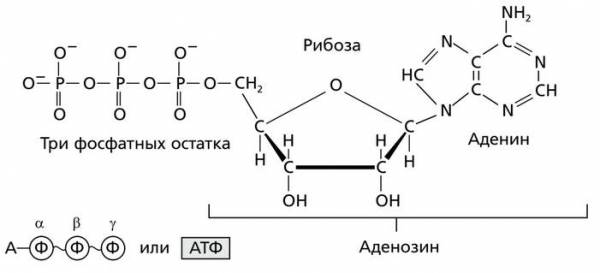 Рибонуклеозидтрифосфат — АТФ (аденозинтрифосфат)
Рибонуклеозидтрифосфат — АТФ (аденозинтрифосфат)
Процесс удвоения ДНК, то есть репликация, начинается с того, что двойная спираль ДНК раскручивается, а затем в определенной точке (или в нескольких точках) обе цепи под действием фермента расходятся под определенным углом, образуя активный участок Y-образной формы, перемещающийся вдоль родительской спирали ДНК, который называют репликационной вилкой. В каждом таком участке ДНК-полимераза осуществляет синтез двух новых дочерних цепей ДНК только в направлении от 5′-конца к 3′-концу (5′ → 3′).
Каждая одинарная цепь присоединяет к себе свободные нуклеотиды, имеющиеся в клетке, и достраивается по принципу комплементарности (А–Т и Г– Ц) до двойной. Таким образом, каждая полинуклеотидная цепочка, выполняя роль шаблона, или матрицы, создает новую цепь. Матрицей (от лат. mater – основа) называют зеркальную основу печатной формы, служащей для получения стереотипных копий. По принципу «копирования с матрицы» осуществляется и репликация дочерних молекул ДНК. В итоге вместо одной молекулы ДНК воссоздаются две молекулы точно такого же нуклеотидного состава, как и первоначальная.
Во время репликации ДНК каждая из образовавшихся двухцепочечных молекул имеет одну нить от материнской ДНК, а другую – вновь образованную – дочернюю молекулу ДНК. Сохранение одной первоначальной (материнской) нити в структуре новой молекулы ДНК получило название полуконсервативности ДНК. Вновь синтезированная молекула ДНК полностью идентична первоначальной.
Обычно репликация начинается одновременно во многих точках двухцепочечной молекулы ДНК. Место начала репликации называют точкой инициации. Длинная цепь ДНК реплицируется не вся сразу, а фрагментами. Участок между двумя точками инициации, в которых осуществляется синтез дочерних нитей, называют репликоном. Репликон является единицей репликации.
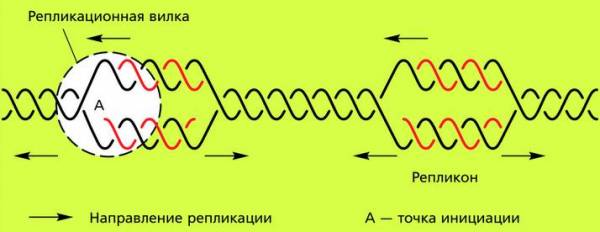 Репликон ДНК эукариотической клетки
Репликон ДНК эукариотической клетки
В каждой молекуле ДНК обычно функционируют несколько репликонов. В каждом репликоне под действием перемещающегося фермента ДНК-полимеразы обе цепи расходятся, образуя вилку, разошедшиеся участки молекулы начинают выполнять роль матрицы, на которой происходит самоудвоение (репликация) ДНК.
Репликационная вилка прекращает свое движение, только когда встречает соседнюю вилку, движущуюся в противоположном направлении.
Процесс самоудвоения
В конце 1950-х годов считалось, что с помощью фермента ДНК-полимеразы каждая отделившаяся цепочка по мере перемещения репликационной вилки от одного конца к другому достраивается на основе комплементарности до новой – дочерней двойной цепи ДНК. Однако дальнейшие исследования показали, что процесс репликации не идет так просто, поскольку две цепи в молекуле ДНК антипараллельны, а ДНК-полимераза ведет сборку биополимера только в направлении от 5′-конца к 3′-концу. Тогда можно ожидать, что рост дочерних молекул по одной цепочке будет происходить в направлении от 5′-конца к З’-концу, а в направлении 3′ → 5′ идти не будет. Следовательно, таким путем, как полагали в 60-е годы XX века, самоудвоение молекулы происходить не может. То, как на антипараллельной цепочке ДНК образуется дочерняя молекула, установили только в начале 70-х годов.
В настоящее время установлено, что если на одной полимерной цепи молекулы ДНК сборка дочерней спирали (направление 5′ → 3′) идет непрерывно и она постепенно удлиняется за счет добавления нуклеотидов на З’-конце, то синтез второй дочерней молекулы на антипараллельной материнской цепи тоже идет в направлении 5′ → 3′, но прерывисто и с заметным отставанием от первой. Поэтому первую, непрерывно образующуюся цепь называют ведущей, или лидирующей, а вторую – отстающей, или запаздывающей.
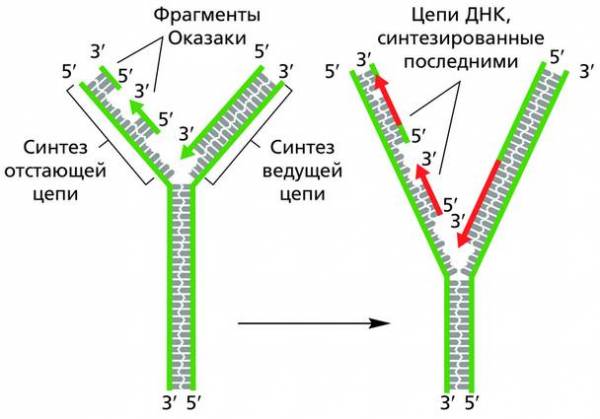 Репликационная вилка и образование дочерних молекул ДНК
Репликационная вилка и образование дочерних молекул ДНК
Заметим, что из-за антипараллельности молекул ДНК образующаяся репликационная вилка оказывается асимметричной. Из двух синтезируемых дочерних цепей одна (ведущая) строится непрерывно и достаточно быстро в направлении 5′ → 3′. Другая – прерывисто и из небольших фрагментов (фрагментов Оказаки), каждый из которых тоже наращивается в направлении 5′ → 3′, хотя в целом вся эта цепь строится на матрице от З’- к 5′-концу. Фрагментарный способ удвоения молекулы занимает больше времени, поэтому данный процесс идет с отставанием в сравнении с репликацией первой цепочки.
Из клеток живых организмов выделено несколько ДНК-полимераз, и в разных лабораториях они получили различные наименования.
После открытия в 1958 году Артуром Корнбергом у Escherichia coli фермента, катализирующего биосинтез ДНК и названного ДНК-полимеразой I, в течение почти 10 лет считалось, что этот фермент является единственной полимеразой, принимающей участие в репликации ДНК in vitro. Но в дальнейшем оказалось, что для репликации ДНК необходимо участие нескольких ферментов. ДНК-полимераза I не наделена способностью инициировать синтез цепей ДНК. Одним из хорошо изученных ферментов, участвующих в стадии инициации репликации ДНК, является специфическая клеточная РНК-полимераза, названная праймазой, которая катализирует синтез первого короткого олигорибонуклеотида (от 10 до 60 нуклеотидов), то есть праймера, с которого затем начинается синтез ДНК.
Основным ферментом, катализирующим биосинтез новообразованной ДНК (ведущей и отстающей цепей), является ДНК-полимераза III, представляющая собой комплекс собственно ДНК-полимеразы (молекулярная масса около 900 тыс.) и ряда других белков.
Этапы биосинтеза ДНК
Основываясь главным образом на данных, полученных в опытах in vitro, предполагают, что условно механизм синтеза ДНК может быть подразделен на три этапа; инициацию, то есть начало, элонгацию – продолжение и терминацию – завершение (прекращение) синтеза. Каждый из этих этапов требует участия специфических ферментов и белковых факторов.
Этап I – инициация биосинтеза ДНК – является началом синтеза дочерних нуклеотидных цепей. В инициации участвует минимум восемь хорошо изученных ферментов и белков. Инициация – единственная стадия репликации ДНК, которая весьма тонко и точно регулируется, однако детальные механизмы ее до сих пор не раскрыты и в настоящее время интенсивно исследуются.
Этап II – элонгация синтеза ДНК – включает два кажущихся одинаковыми, но резко различающихся по механизму синтеза лидирующей и отстающей цепей на обеих материнских цепях ДНК. Синтез лидирующей цепи начинается с синтеза праймера (при участии праймазы) у точки начала репликации, затем к праймеру присоединяются дезоксирибонуклеотиды под действием ДНК-полимеразы III. Далее синтез протекает непрерывно, следуя шагу репликационной вилки. Синтез отстающей цепи, напротив, протекает в направлении, обратном движению репликационной вилки, и происходит фрагментарно. Фрагменты всякий раз синтезируются раздельно, начиная с синтеза праймера, который может переноситься с помощью одного из белковых факторов репликации с готового фрагмента в точку старта биосинтеза последующего фрагмента противоположно направлению синтеза дочерней цепи (подобно шитью иголкой назад). На этом участке сборка фрагментов идет при участии ДНК-полимеразы, но в направлении 5’→ 3′ (встречно). Элонгация завершается заполнением (достраиванием) освободившихся мест (брешей) комплементарными дезоксирибонуклеотидами под действием той же праймазы, объединением образовавшихся небольших фрагментов ДНК (их называют фрагментами Оказаки, по имени ученого, открывшего это явление) с помощью ДНК-лигаз и формированием дочерней цепи ДНК.
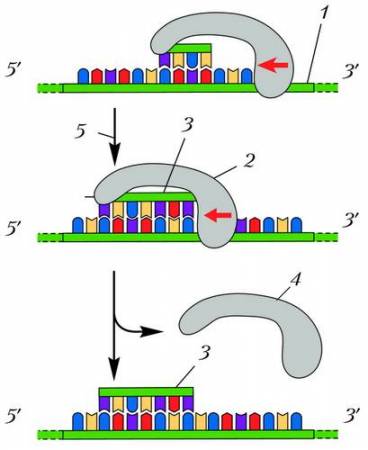 Схема сборки фрагментов отстающей цепи ДНК: 1 – антипараллельная цепь ДНК (матрица); 2 – фермент РНК-праймаза переносит начало сборки нового фрагмента; 3 – участок, собранный с помощью ДНК-полимеразы; 4 – переход праймазы на новый участок синтеза; 5 – начало синтеза дочерней ДНК
Схема сборки фрагментов отстающей цепи ДНК: 1 – антипараллельная цепь ДНК (матрица); 2 – фермент РНК-праймаза переносит начало сборки нового фрагмента; 3 – участок, собранный с помощью ДНК-полимеразы; 4 – переход праймазы на новый участок синтеза; 5 – начало синтеза дочерней ДНК
Этап III – терминация синтеза ДНК – наступает, скорее всего, когда исчерпана ДНК-матрица и трансферазные реакции (реакции переноса) прекращаются. Точность репликации ДНК чрезвычайно высока, возможна одна ошибка на 1010 трансферазных реакций, однако подобная ошибка обычоо легко исправляется за счет процессов репарации (восстановления).
Функцию раскручивания (расплетения) двойной спирали ДНК в репликационной вилке, происходящего за счет энергии гидролиза АТФ, выполняет специфический белок, названный геликазой (молекулярная масса 300 тыс.). Образовавшиеся на определенное время одноцепочечные участки ДНК служат в качестве матрицы при репликации и стабилизируются при помощи особых белков, связывающихся с одноцепочечной ДНК (ДНК-связывающие белки) и препятствующих обратному комплементарному взаимодействию цепей ДНК (молекулярная масса 75 600). В связи с этим их иногда называют дестабилизирующими.
Дестабилизирующие белки (1) и фермент ДНК-геликаза (2) обеспечивают деспирализованность цепи ДНК в репликационной вилке
Мы привели лишь схематическое описание процесса репликации ДНК. На самом деле этот процесс очень сложен, в нем участвует множество специфических белков и различных ферментов, способствующих расплетению ДНК и предотвращению ее спутывания, скручивания, а также обеспечивающих сшивание фрагментов ДНК в целостную двойную спираль.
Имеются, кроме того, особые ферменты топоизомеразы (у прокариот одна из них названа ДНК-гиразой), которые играют особую роль в сверхспирализации, обеспечивая как репликацию, так и транскрипцию ДНК. Эти ферменты наделены способностью не только создавать супервитки, но и уничтожать суперспирализацию путем сшивания образующихся разрывов или разрезания ДНК. Наконец, открыты специальные ферменты, «редактирующие» ДНК, то есть осуществляющие вырезание и удаление ошибочно включенных нуклеотидов или репарирующие повреждения ДНК, вызванные физическими или химическими факторами (рентгеновское излучение, УФ-лучи, химический мутагенез и др.).
Из приведенного неполного перечня участников репликации ДНК можно понять, каким образом осуществляется этот процесс и какова его сложность.
Таким образом, процесс репликации ДНК характеризуется рядом принципиальных особенностей. Среди них: 1) комплементарность оснований и матричный характер синтеза; 2) полуконсервативность; 3) антипараллельность; 4) прерывистость синтеза; 5) точность копирования генетического кода; б) полная идентичность дочерних молекул материнской молекуле ДНК.