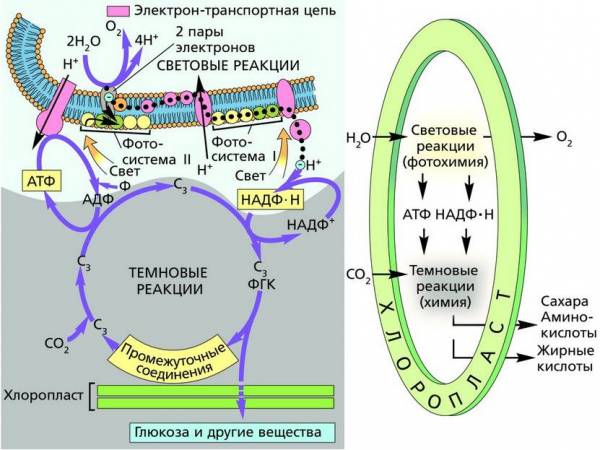
Закон независимого наследования генов демонстрирует дискретность генетического материала. Это проявляется в независимом комбинировании аллелей разных генов, в их фенотипическом выражении. Однако в ряде случаев идентификация фенов (проявлений элементарных признаков) сопряжена с некоторыми трудностями.
Дело в том, что генотип любого организма представляет собой систему взаимодействующих между собой генов. Некоторые признаки организма определяются действием аллелей не одного, а одновременно нескольких генов. Выявить это взаимодействие генов позволяет генетический анализ на основе изучения расщепления по фенотипу.
Простейшие формы взаимодействия аллельных генов – отношения доминирования – происходят в соответствии с законами Менделя. Однако известно еще несколько типов взаимодействия генов между собой, которые приводят к отклонению частоты встречаемости признаков от ожидаемой согласно законам Менделя. Это наблюдается при взаимодействии аллельных и неаллельных генов. Рассмотрим некоторые формы этого взаимодействия.
СОДЕРЖАНИЕ
Типы взаимодействия аллельных генов
При взаимодействии аллелей одного гена различают полное доминирование, неполное доминирование, кодоминирование и множественный аллелизм.
При полном доминировании действие одной аллели полностью подавляет действие другой аллели по типу доминантного и рецессивного взаимодействия. Неполное доминирование свидетельствует о промежуточном характере выражения признака у гетерозигот. Пример такого наследования можно наблюдать у цветкового растения ночная красавица (Mirabilis jalapa). У гибридов первого поколения (F1) все гетерозиготные особи имеют розовые цветки, а у F2 наблюдается расщепление в соотношении 1AA (красные цветки) : 2Aa (розовые цветки) : 1aa (белые цветки).
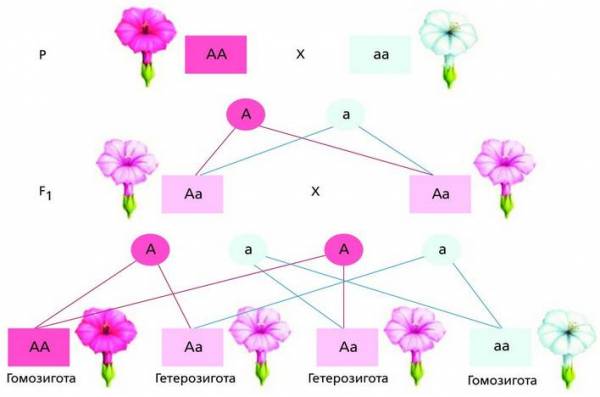 Наследование окраски цветков у ночной красавицы при неполном доминировании Кодоминирование (от лат. co – с, вместе и доминирование) характеризует взаимодействие разных аллелей одного гена, в котором каждая из них имеет самостоятельное проявление. В данном случае наблюдается совместное участие обеих аллелей в определении признака у гетерозиготной особи как частный случай доминантности. Классическим примером кодоминирования служит взаимодействие аллелей, определяющих группы крови в системе AB0, когда два доминирующих гена вместе, не подавляя друг друга, определяют признак (группу крови AB).
Наследование окраски цветков у ночной красавицы при неполном доминировании Кодоминирование (от лат. co – с, вместе и доминирование) характеризует взаимодействие разных аллелей одного гена, в котором каждая из них имеет самостоятельное проявление. В данном случае наблюдается совместное участие обеих аллелей в определении признака у гетерозиготной особи как частный случай доминантности. Классическим примером кодоминирования служит взаимодействие аллелей, определяющих группы крови в системе AB0, когда два доминирующих гена вместе, не подавляя друг друга, определяют признак (группу крови AB).
У гена I известны три типа аллелей (I A , I B , I 0 ), детерминирующих группы крови A, B, AB и 0. Группа крови A определяется гомозиготным состоянием I A I A , при котором эритроциты имеют только поверхностный антиген A. При гомозиготности по другой аллели – I B I B – на мембране эритроцитов присутствует другой антиген – B (группа крови B). В случае гомозиготности по аллели I 0 I 0 эритроциты лишены A- и B-антигенов (группа крови 0). В случае гетерозиготности I A I 0 или I B I 0 группа крови будет соответственно или A, или B, то есть происходит полное доминирование. Если же человек гетерозиготен одновременно по двум аллелям I A и I B , то наблюдается случай кодоминирования: эритроциты несут оба антигена (группа крови AB). Аллели I A и I B работают в гетерозиготе как бы независимо друг от друга, что определяется с помощью иммунологических методов.
Множественный аллелизм. В популяциях нередко встречаются гены, имеющие не две аллели, а значительно большее их количество. Это явление называют множественным аллелизмом. Таким способом детерминируются, например, окраска шерсти животных, окраска глаз у дрозофилы, форма белых пятен на листьях клевера ползучего и др. Наследование групп крови у человека также иллюстрирует проявление множественного аллелизма, поскольку ген I может быть представлен тремя разными аллелями (I A , I B , I 0 ). Эти аллели комбинируются в зиготах только попарно.
Типы взаимодействия неаллельных генов
К нарушению закономерностей расщепления по Менделю приводит и взаимодействие неаллельных генов; комплементарность (от лат. complementum – дополнение), эпистаз (от греч. epistasis – остановка, препятствие), полимерия (от греч. polymeria – множественность), плейотропия (от греч. pleion – многочисленный и tropos – направление) и действие генов-модификаторов (от лат. modus – мера, вид и facio – делаю).
Комплементарность
Комплементарность свидетельствует о развитии нового проявления признака, отличного от родительских вариантов, появившегося в связи с взаимодействием неаллельных генов. Комплементарное взаимодействие аллелей (доминантных или рецессивных) обычно рассматривается при дигибридном скрещивании.
Например, у плодовой мушки дрозофилы имеются две рецессивно наследуемые мутации: ярко-красные глаза (scarlet — st) и коричневые глаза (brown — bw). Доминантную аллель обозначают знаком «плюс» (+). При скрещивании особей с ярко-красными (stst) (один признак) и коричневыми (bwbw) (второй признак) глазами у всех гибридов первого поколения темно-красные глаза (появляется третий признак, которого не было у родителей). При скрещивании между собой гибридов первого поколения во втором поколении (F2) наблюдается расщепление в соотношении 9 (темно-красные глаза) : 3 (коричневые глаза) : 3 (ярко-красные глаза) : 1 (белые глаза).
Результаты скрещивания дрозофил с ярко-красными и коричневыми глазами
Таким образом, в первом поколении проявляется признак, отличный от родительских вариантов, – темно-красные глаза, что свидетельствует о взаимодействии генов. Появление же во втором поколении (F2) четырех, а не двух, как при моногибридном скрещивании, фенотипических классов указывает на дигибридное скрещивание с сохранением полного доминирования по обоим признакам. Поэтому появление в F1 мушек с темно-красными глазами является следствием взаимодействия доминантных аллелей генов (st+ и bw+). Появление во втором поколении мушек с новым признаком (белые глаза) – следствие взаимодействия рецессивных аллелей этих же генов (stst и bwbw).
Биохимический анализ позволяет дополнить эти результаты скрещивания мух. Известно, что у дрозофилы окраска глаз обусловлена двумя пигментами – красным и бурым. Рецессивная аллель bw в гомозиготе прерывает синтез красного пигмента, поэтому глаза мух содержат только бурый пигмент. Рецессивная аллель st в гомозиготе блокирует синтез бурого пигмента, поэтому в глазах мух присутствует только красный пигмент. Когда в дигетерозиготе оказываются нормальные (доминантные) аллели обоих генов, то синтезируются оба пигмента. Результат в F1 – комплементарное взаимодействие нормальных аллелей. Если в F2 в гомозиготе оказывается и bwbw, и stst, то не синтезируются ни красный, ни бурый пигменты и глаза оказываются белыми, неокрашенными.
По типу комплементарности взаимодействуют также гены, определяющие, например, форму плода тыквы.
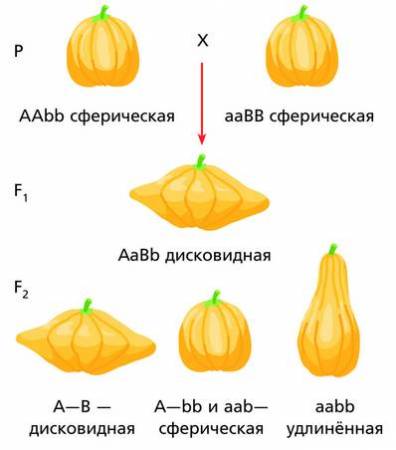 Комплементарное взаимодействие генов, определяющих форму плода тыквы (соотношение 9 : 6 : 1)
Комплементарное взаимодействие генов, определяющих форму плода тыквы (соотношение 9 : 6 : 1)
Эпистаз
Эпистаз – одна из форм взаимодействия генов, при которой аллели одного гена подавляют проявление аллелей других генов, например A > B или A > b. Такие гены-подавители называют эпистатическими генами, ингибиторами или супрессорами. Подавляемые гены называяют гипостатическими. Примером может служить взаимодействие генов bw и st у дрозофилы, рассмотренное выше. Соотношение фенотипов в F2 является следствием того, что рецессивная аллель bw в гомозиготе препятствует проявлению доминантной аллели st+, а рецессивная аллель st подавляет проявление bw+.
Полимерия
Существует множество примеров, показывающих, что степень развития одного и того же признака обусловлена влиянием целого ряда неаллельных генов, проявляющихся сходным образом. Все они как бы дублируют действие друг друга в проявлении данного признака. Это явление множественного действия генов называют полимерией, а сами гены – полимерами. Их обозначают одними и теми же буквенными символами: A1, a1 – доминантная и рецессивная аллели первого гена; A2, a2 – второго гена и т. д.
По типу полимерии наследуются цвет кожи человека, высота растений, количество белка в эндосперме семян, скорость роста и масса тела животных, яйценоскость кур, молочность коров, длина колоса у злаков, содержание сахара в корнеплодах сахарной свеклы и многие другие количественные признаки организма. Количественные признаки могут усиливаться за счет применения «кумулятивной полимерии», охватывающей несколько полимерных генов. При этом интенсивность действия кумулятивной полимерии может усиливаться пропорционально числу доминантных аллелей разных генов.
Плейотропия
Плейотропия – это множественное действие генов, когда один ген определяет развитие одновременно не одного, а сразу нескольких признаков организма. Например, у некоторых цветковых растений гены, контролирующие красную (антоциановую) окраску цветка, одновременно контролируют и красную окраску стебля. У человека известен доминантный ген, определяющий аномально длинные и тонкие пальцы кистей и стоп – «паучьи пальцы» (синдром Марфана). Этот же ген определяет и аномалии хрусталика глаза, и порок сердца. Взаимодействие совершается на уровне продуктов реакций, контролируемых генами.
Действие генов-модификаторов
Наряду с генами «основного» действия на развитие любого признака влияют другие, зачастую неизвестные нам гены. Эти гены, получившие название модификаторов, не определяют какую-либо конкретную реакцию или развитие признака, но способны усиливать или ослаблять, то есть модифицировать, проявление действия «основных», или «главных», генов. Многие гены одновременно могут быть «основными» по одним признакам и модификаторами по другим.
Таким образом, изучение взаимодействия генов (аллельных и неаллельных) показало, что любой наследственный признак определяется многими генами, точнее, всем генотипом. Это свидетельствует о том, что генотип является не суммой генов, а сложной целостной системой взаимодействующих генов, определяющей наследственность организма.