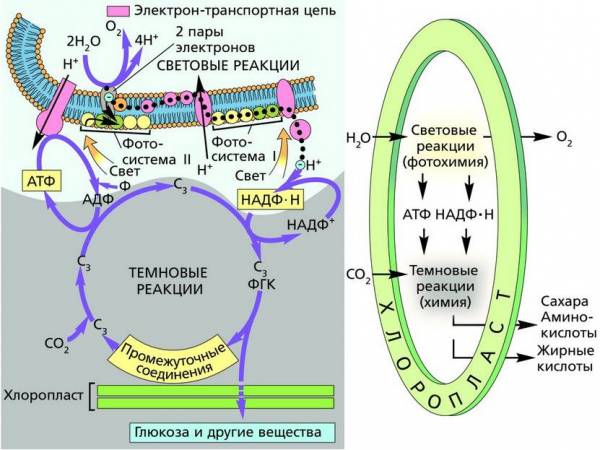
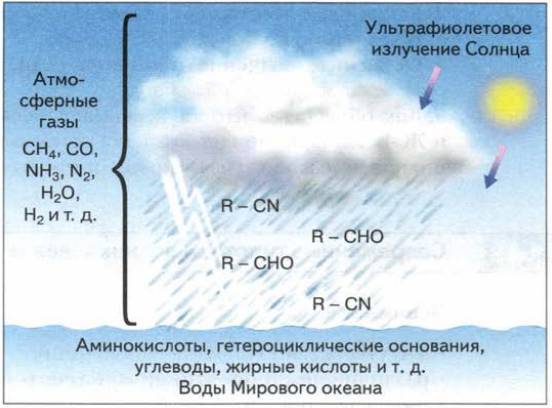
Эукариоты, возможно, существуют на Земле около 2,7 млрд лет. Эта дата определена благодаря обнаружению в древних отложениях остатков стероидных соединений, встречающихся только в мембранах эукариотических клеток. Однако только примерно 1–1,5 млрд лет назад эукариоты стали превалирующим компонентом биосферы, потеснив прокариот. Появление эукариот – событие планетарного масштаба, так как именно они дали начало огромному многообразию организмов, в том числе ландшафтообразующих. Не случайно этот период в истории Земли называют неопротерозойской революцией.
Гипотезы о происхождении эукариот
С момента обнаружения существенных различий в строении клеток бактерий и простейших, в первую очередь отсутствия или присутствия ядра, встал вопрос о выяснении происхождения и степени родства этих групп. Были выдвинуты три основные гипотезы происхождения эукариот и их органоидов – аутогенная, химерная и симбиогенная, которые и в настоящее время все еще остаются гипотезами.
Согласно аутогенной (от греч. autos – сам, собственный и genesis – происхождение, возникновение), или сукцессионной, гипотезе эукариотическая клетка самостоятельно развилась из прокариотической в результате впячивания плазматической мембраны с ее последующей перестройкой. Таким образом, все мембранные органоиды, в том числе митохондрии и пластиды, ведут свое начало от имеющихся у прокариот внутриплазматических мембранных структур, подобных мезосомам. В настоящее время аутогенная гипотеза считается устаревшей.
Химерная гипотеза трактует образование клетки эукариот путем слияния целых клеток различных прокариот. Предложено несколько вариантов химерного происхождения эукариотической клетки от архей: слияние с фототрофной протеобактерией; слияние архебактерии с эубактерей-бродильщиком, обладающей гликолизом; слияние архебактерии со спирохетой, приведшее к образованию жгутика.
Согласно симбиогенной гипотезе, или гипотезе симбиогенеза, первичная клетка крупной прокариотической бактерии вступала в симбиоз с другими прокариотами и в результате постепенно приобретала различные органоиды: из симбионтных цианобактерий образовались пластиды, из гетеротрофных бактерий – митохондрии, из каких-то иных симбионтов – центриоли и жгутики.
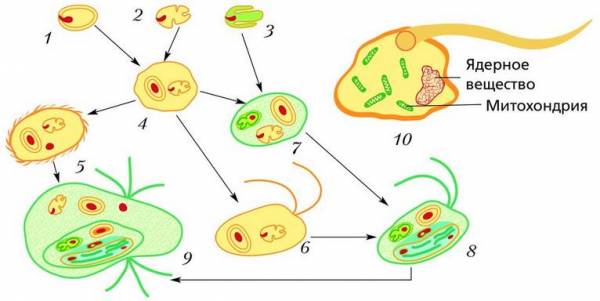 Схема возникновения эукариот согласно симбиогенной гипотезе: 1 – прокариота – будущее ядро; 2 – прокариота – будущая митохондрия; 3 – прокариота – будущий хлоропласт; 4 – первая эукариота с ядром и митохондрией; 5–9 – возможные пути дальнейшего усложнения структуры клетки эукариот; 10 – бактерия-симбионт с митохондриями (первая эукариотическая клетка)
Схема возникновения эукариот согласно симбиогенной гипотезе: 1 – прокариота – будущее ядро; 2 – прокариота – будущая митохондрия; 3 – прокариота – будущий хлоропласт; 4 – первая эукариота с ядром и митохондрией; 5–9 – возможные пути дальнейшего усложнения структуры клетки эукариот; 10 – бактерия-симбионт с митохондриями (первая эукариотическая клетка)
Некоторые сторонники этой гипотезы предполагали даже возможность симбиотического происхождения ядра.
В первую очередь именно сходство митохондрий и хлоропластов с прокариотическими клетками натолкнуло ученых на мысль об эндобионтном (от греч. endon – внутри и biontos – живущий) происхождении некоторых органоидов. При этом предполагается, что прокариоты были внутриклеточными «жильцами» клеток эукариот. Возможно, в глубокой древности некие прокариотические клетки, способные к значительному производству АТФ, поселялись в других клетках и питались за их счет. Первоначально они были паразитами, однако позже клетка-хозяин начала использовать производимую ими АТФ для собственных нужд. Возможно также, что какие-то древние фотосинтезирующие прокариоты, ведя внутриклеточный симбиотический образ жизни с клеткой-хозяином, взаимовыгодно снабжали друг друга различными продуктами своего метаболизма. В процессе эволюции эти связи закрепились, и симбиотические прокариотные организмы, утратив самостоятельность, превратились в органоиды клеток эукариот.
Все эти предположения породили ряд гипотез, объясняющих вероятность подобных явлений. Но в настоящее время ученые еще так и не пришли к единому мнению о происхождении многих внутриклеточных структур эукариот. Однако сама дискуссия вызвала массу интересных работ по изучению митохондрий и пластид, что обеспечило более глубокое и детальное выяснение свойств этих органоидов и других внутриклеточных структур.
Следует отметить, что впервые идею о симбиотическом происхождении сложно устроенной клетки высказал в конце 60-х годов XIX века русский ученый-ботаник Андрей Сергеевич Фаминцын (1835–1918) на основе изучения лишайников. В начале XX века (1905–1909) биолог Константин Сергеевич Мережковский (1855–1921) выдвинул идею о симбиотическом происхождении носителей пигментов в растительных клетках, которая в 1921 – 1924 годах ботаником Борисом Михайловичем Козо-Полянским (1890– 1957) была развита в симбиотическую гипотезу происхождения клеток растений и животных. В 1983 году сотрудницей Массачусетского университета Линн Маргулис (Саган) гипотеза симбиогенеза была изложена в виде целостной концепции. В последующие годы благодаря новым методам исследований и открытиям симбиогенная гипотеза получила многочисленные подтверждения, и иногда ее называют уже не гипотезой, а теорией.
Гипотезы симбиогенеза по-разному отображают и начало самого процесса, и путь возникновения сложных клеток эукариот.
В настоящее время общепризнанной считается гипотеза симбиотического происхождения эукариотической клетки от прокариоты, вероятнее всего – от архебактерии, хотя не исключено, что предком эукариот была вымершая группа каких-то прокариот, одинаково далекая и от эубактерий, и от архебактерий.
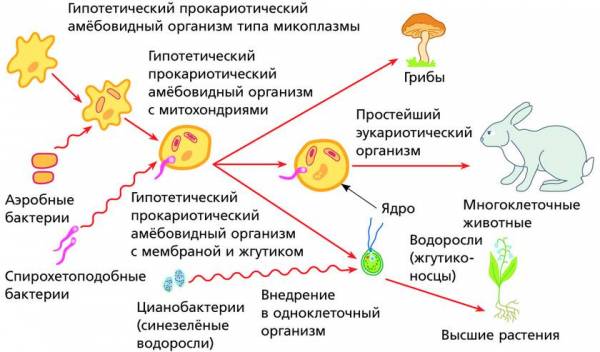 Схема возможного происхождения эукариот от гипотетического прокариотического организма
Схема возможного происхождения эукариот от гипотетического прокариотического организма
Получено достаточное количество данных, доказывающих, что митохондрии происходят от симбиотических альфапротеобактерий (близких к современным пурпурным бактериям), а пластиды растений и большинства водорослей – от симбиотических фотосинтезирующих цианобактерий (синезеленых водорослей). Однако до сих пор не прекращаются дискуссии по поводу природы клетки-хозяина, и главное – относительно происхождения ядра.
Например, согласно одной из гипотез образование ядерной мембраны могло стать побочным результатом избыточной мембранообразующей деятельности эубактерий, поселившихся в цитоплазме архебактерий. Согласно другой гипотезе клеточное ядро сформировалось у архей в результате вирусной инфекции. Существует множество других гипотез возникновения ядра, но ни одна пока не доказана.
Один из наиболее вероятных вариантов происхождения ядра – появление его вследствие впячивания участков плазматической мембраны с находящимися на них точками прикрепления хромосом внутрь полиплоидной (имеющей несколько хромосом) прокариотной клетки. Таким путем первичные эукариоты могли получить ядро, ограниченное двойной ядерной оболочкой, но у них еще сохранялось кольцевое строение хромосом и отсутствовали гистоны. В какой-то мере этот эволюционный путь подтверждается тем, что в ядре большинства эукариот хромосомы также прикрепляются к субмембранным структурам внутренней ядерной мембраны и в ядре всегда имеются плазмиды (внехромосомные молекулы ДНК, часто представляющие собой кольцевые двухцепочечные структуры), а у динофлагеллят (панцирных жгутиконосцев) хромосомы содержат кольцевые молекулы ДНК, практически лишенные гистонов.
Имеется предположение о том, что наследственный материал от бактерий мог быть получен в результате горизонтального переноса генов, например при участии плазмид или вирусов, а также при половом процессе с неродственными организмами или при химерном слиянии клеток. Возможно также, что предковая архея растворяла оболочки бактерий, «высасывала» их содержимое, в том числе ДНК, а затем встраивала эту ДНК в свой геном. Для регуляции массивного химерного генетического аппарата появились новые, более сложные системы считывания, стабилизации ДНК и др.
Как видим, гипотез много и они касаются многих структур эукариотической клетки, но в целом все они поддерживают главную идею симбиогенеза: от прокариот – к эукариотам.