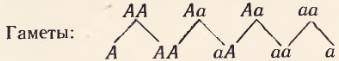
Сцепленное наследование. Независимое распределение генов (второй закон Менделя) основано на том, что гены, относящиеся к разным аллелям, размещены в разных парах гомологичных хромосом. Естественно возникает вопрос: а как же будет происходить распределение разных (неаллельных) генов в ряду поколений, если они лежат в одной и той же паре хромосом? Такое явление должно иметь место, ибо число генов во много раз превосходит число хромосом. Очевидно, к генам, находящимся в одной хромосоме, закон независимого распределения (второй закон Менделя) не применим. Он ограничен лишь теми случаями, когда гены разных аллелей находятся в различных хромосомах.
Закономерность наследования при нахождении генов в одной хромосоме была тщательно изучена Т. Морганом и его школой Основным объектом исследований служила небольшая плодовая мушка дрозофила 108. Это насекомое исключительно удобно для генетической работы. Мушка легко разводится в лабораторных условиях, плодовита, каждые 10-15 дней при оптимальной для нее температуре 25-26°С дает новое поколение, обладает многочисленными и разнообразными наследственными признаками, имеет небольшое число хромосом (в диплоидном наборе –  .
.
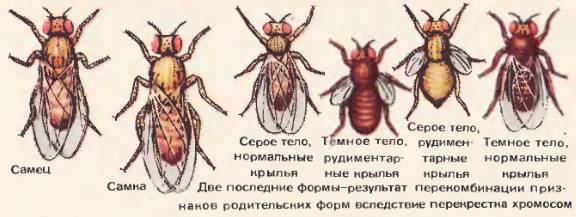 Рис. 108. Разные наследственные формы мухи дрозофилы
Рис. 108. Разные наследственные формы мухи дрозофилы
Опыты показали, что гены, локализованные в одной хромосоме, оказываются сцепленными, т. е. наследуются преимущественно вместе, не обнаруживая независимого распределения. Рассмотрим конкретный пример. Если скрестить дрозофилу с серым телом и нормальными крыльями с мушкой, обладающей темной окраской тела и зачаточными крыльями, то в первом поколении гибридов все мушки будут серыми, с нормальными крыльями, Это гетерозигота по двум парам аллелей (серое тело темное тело и нормальные крылья – зачаточные крылья). Проведем скрещивание. Скрестим самок этих дигетерозиготных мух (серое тело и нормальные крылья) с самцами, обладающими рецессивными признаками – темным телом и зачаточными крыльями. Исходя из второго закона Менделя, можно было бы ожидать получения в потомстве мух четырех фенотипов: 25% серых, с нормальными крыльями; 25% серых, с зачаточными крыльями; 25% темных, с нормальными крыльями; 25% темных, с зачаточными крыльями.
На самом деле в опыте мух с исходной комбинацией признаков (серое тело – нормальные крылья, темное тело – зачаточные крылья) оказывается значительно больше (в данном опыте по 41,5%), чем мух с перекомбинированными признаками (серое тело – зачаточные крылья и темное тело – нормальные крылья). Их будет всего по 8,5% каждого типа [108]. На этом примере видно, что гены, обусловившие признаки серое тело – нормальные крылья и темное тело – зачаточные крылья, наследуются преимущественно вместе, или, иначе говоря, оказываются сцепленными между собой. Это сцепление является следствием локализации генов в одной и той же хромосоме. Поэтому при мейозе эти гены не расходятся, а наследуются вместе. Явление сцепления генов, локализованных в одной хромосоме, известно под названием закона Моргана.
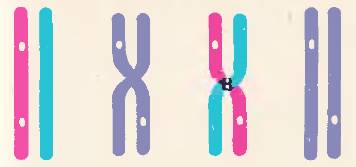 Рис. 109. Схема перекреста хромосом
Рис. 109. Схема перекреста хромосом
Два гена, расположенных в одной хромосоме (светлые кружки в красной хромосоме) в результате перекреста оказываются в разных гомологичных хромосомах
Почему же все-таки среди гибридов второго поколения появляется небольшое число особей с перекомбинацией родительских признаков? Почему сцепление генов не является абсолютным? Исследования показали, что эта перекомбинация генов обусловлена тем, что в процессе мейоза при конъюгации гомологичных хромосом они иногда обмениваются своими участками, или, иначе говоря, между ними происходит перекрест [109], что при этом гены, находившиеся первоначально в одной из двух гомологичных хромосом, окажутся в разных гомологичных хромосомах. Между ними произойдет перекомбинация. Частота перекреста для разных генов оказывается различной. Это зависит от расстояния между ними. Чем ближе в хромосоме расположены гены, тем реже они разделяются при перекресте. Это происходит потому, что хромосомы обмениваются различными участками, и близко расположенные гены имеют больше вероятности оказаться вместе. Исходя из этой закономерности удалось для хорошо изученных в генетическом отношении организмов построить генетические карты хромосом, на которых нанесено относительное расстояние между генами.
Биологическое значение перекреста хромосом очень велико. Благодаря ему создаются новые наследственные комбинации генов, повышается наследственная изменчивость, которая поставляет материал для естественного отбора.
Генетика пола. Хорошо известно, что у раздельнополых организмов (в том числе и у человека) соотношение полов обычно составляет 1:1. Какие причины определяют пол развивающегося организма? Вопрос этот издавна интересовал человечество ввиду его большого теоретического и практического значения. Хромосомный набор самцов и самок у большинства раздельнополых организмов неодинаков. Познакомимся с этими различиями на примере набора хромосом у дрозофилы [110]. По трем парам хромосом самцы и самки не отличаются друг от друга. Но в отношении одной пары имеются существенные различия. У самки две одинаковые (парные) палочковидные хромосомы; у самца только одна такая хромосома, пару которой составляет особая, двуплечая хромосома. Те хромосомы, в отношении которых между самцами и самками нет различий, называют аутосомами. Хромосомы, по которым самцы и самки отличаются друг от друга, называют половыми. Таким образом, хромосомный набор дрозофилы слагается из шести аутосом и двух половых хромосом. Половую, палочковидную хромосому, присутствующую у самки в двойном числе, а у самца – в единичном, называют X-хромосомой; вторую, половую (двуплечую хромосому самца, отсутствующую у самки) – Y-хромосомой.
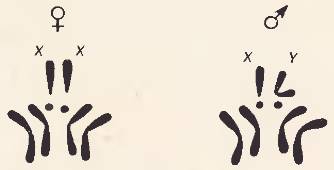 Рис. 110. Хромосомные комплексы самки и самца дрозофилы
Рис. 110. Хромосомные комплексы самки и самца дрозофилы
Каким образом рассмотренные половые различия в хромосомных наборах самцов и самок поддерживаются в процессе размножения? Для ответа на этот вопрос необходимо выяснить поведение хромосом в мейозе и при оплодотворении. Сущность этого процесса представлена на рисунке 111. При созревании половых клеток у самки каждая яйцеклетка в результате мейоза получает гаплоидный набор из четырех хромосом: три аутосомы и одну X-хромосому. У самцов в равных количествах образуются сперматозоиды двух сортов. Одни несут три аутосомы и X-хромосому, другие – три аутосомы и Y-хромосому. При оплодотворении возможны две комбинации. Яйцеклетка с равной вероятностью может быть оплодотворена спермием с X- или Y-хромосомой. В первом случае из оплодотворенного яйца разовьется самка, а во втором – самец. Пол организма определяется в момент оплодотворения и зависит от хромосомного набора зиготы.
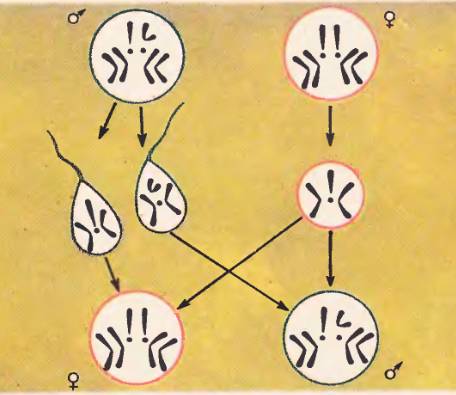 Рис. 111. Механизм определения пола у дрозофил
Рис. 111. Механизм определения пола у дрозофил
У самца образуются гаметы двух категорий: одни несут в гаплоидном наборе X-хромосомы, другие Y-хромосомы
У человека хромосомный механизм определения пола тот же, что и у дрозофилы. Диплоидное число хромосом человека – 46. В это число входят 22 пары аутосом и 2 половые хромосомы. У женщин это две X-хромосомы, у мужчин – одна X- и одна Y-хромосома [112]. Соответственно у мужчин образуются сперматозоиды двух сортов – с X- и Y-хромосомами.
 Рис. 112. Хромосомный комплекс мужчины (слева) и женщины (справа)
Рис. 112. Хромосомный комплекс мужчины (слева) и женщины (справа)
У некоторых раздельнополых организмов (например, некоторых насекомых) Y-хромосома вообще отсутствует. В этих случаях у самца оказывается на одну хромосому меньше: вместо X- и Y- у него имеется одна X-хромосома. Тогда при образовании мужских гамет в процессе мейоза X-хромосома не имеет партнера для конъюгации и отходит в одну из клеток. В результате половина всех сперматозоидов имеет X-хромосому, а другая половина лишена ее. При оплодотворении яйца спермием с X-хромосомой получается комплекс с двумя X-хромосомами, и из такого яйца развивается самка. Если яйцеклетка будет оплодотворена спермием без X-хромосомы, то разовьется организм с одной X-хромосомой (полученной через яйцеклетку от самки), который будет самцом.
Во всех рассмотренных выше примерах развиваются спермии двух категорий: либо с X- и Y-хромосомами (дрозофила, человек), либо половина спермиев несет X-хромосому, а другая совсем лишена половой хромосомы. Яйцеклетки в отношении половых хромосом все одинаковы. Во всех этих случаях мы имеем мужскую гетерогаметность (разногаметность). Женский пол гомогаметен (равногаметен). Наряду с этим в природе встречается и другой тип определения пола, характеризующийся женской гетерогаметностью. Здесь имеют место отношения обратные только что рассмотренным. Разные половые хромосомы или только одна X-хромосома свойственны женскому полу. Мужской пол обладает парой одинаковых X-хромосом. Очевидно, в этих случаях будет иметь место женская гетерогаметность. После мейоза образуются яйцевые клетки двух сортов, тогда как в отношении хромосомного комплекса все спермии одинаковы (все несут одну X-хромосому). Следовательно, пол зародыша будет определяться тем, какое яйцо – с X- или Y-хромосомой будет оплодотворено.
Женская гетерогаметность имеет место у некоторых насекомых, например у бабочек. Среди позвоночных животных она характерна для птиц и пресмыкающихся.
1. При какой форме взаимного расположения неаллельных генов осуществляется второй закон Менделя (независимого распределения) и при какой – закон сцепления Моргана? 2. В результате какого процесса нарушается сцепление генов, локализованных в одной хромосоме? 3. В чем заключается генетический механизм определения пола?