Живые организмы осуществляют постоянный обмен веществ и энергии с окружающей средой.
В организмах постоянно идут процессы биосинтеза.
Биосинтез — это построение из простых веществ более сложных.
В клетках обнаружено более 1000 ферментов.
Любые реакции, происходящие в клетке, идут с поглощением Q.
Различают первичный метаболизм — синтез и распад белков, НК, липидов, углеводов и аскорбиновой кислоты (характерен для всех живых организмов); и вторичный метаболизм — это образование и превращение других классов органических веществ (характерен для грибов, растений и некоторых прокариот (антибиотики)).
Пластический обмен — это совокупность реакций биологического синтеза. Из веществ, поступающих из вне, образуются соединения подобные веществам клетки.
Биосинтез белка
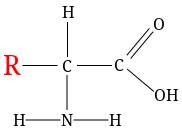
Свойства белка определяются его первичной структурой.
Каждой аминокислоте на молекуле ДНК и РНК соответствует три нуклеотида, т.е. последовательность нуклеотидов ДНК (РНК) — это генетический код, несущий информацию о структуре белка.
Генетический код всех организмов обладает общими свойствами:
- Избыточность
Г, Ц, Т, А: 4 3 = 64 — общее число возможных комбинаций, но кодируется всего 20 аминокислот; т.е. одной аминокислоте соответствует несколько кодов.
Избыточность повышает надежность генетического кода на случай мутаций. - Специфичность . Один триплет всегда соответствует только одной аминокислоте.
- Универсальность . Подобный механизм биосинтеза белка характерен для всех живых организмов — от вируса до человека (ТТТ — отвечает за аминокислоту лизин у всех видов).
- Дискретность . Триплеты не перекрываются. У некоторых вирусов это свойство не всегда проявляется.
- В одной молекуле ДНК записана информация о нескольких сотнях различных белков.
- Гены, отвечающие за синтез белков, в молекуле ДНК разграничены между собой триплетами:
- триплеты-инициаторы начинают синтез и-РНК (АУГ, ГУГ);
- триплеты-терминаторы прекращают синтез и-РНК (УАА, УАГ, УГА).
- И-РНК несет информацию об одном белке.
- Доставка информации к рибосомам, где происходит синтез белка, происходит в два этапа:
a. транскрипция — синтез и-РНК на ДНК;
b. трансляция (от лат. «перенесение»).
У эукариот и-РНК выносится из ядра в цитоплазму. У прокариот синтез белка может начаться еще во время синтеза и-РНК.
У эукариот образуется комплекс между специальными белками и и-РНК, который и переходит в цитоплазму.
В цитоплазме со стороны триплета-инициатора подходит рибосома и начинается синтез полипептида (белка).
Триплеты распознаются т-РНК, которая может переносить только одну аминокислоту.
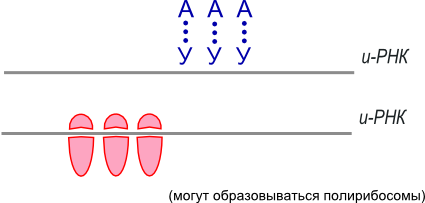
За 1 секунду 1 рибосома способна присоединить 20 аминокислот.
Энергетический обмен (диссимиляция, катаболизм)
При разрыве макроэргической связи выделяется большое количество энергии (АТФ, ГТФ и др.).
Эти вещества обеспечивают в организме проведение импульсов, синтез и распад различных веществ, активный транспорт веществ и др.
В органических соединениях химическая энергия заключена в виде ковалентных связей между атомами (в одном моле глюкозы (180 г) заключено примерно 2800 кДж).
В одной клетке содержится порядка 1 млрд. АТФ.
Энергетический обмен делится на 3 этапа:
- Подготовительный (пищеварение)
Происходит вне клетки. При этом выделяется небольшая часть Q, которая рассеивается в виде тепла.
Полимеры распадаются до мономеров. - Бескислородный
Мономеры поступают в клетку.
a. Гликолиз — бескислородное ферментативное расщепление глюкозы:
C6H12O6 + 2H3PO4 + 2АДФ → 2C3H4O3 + 2H2O + 2АТФ
C3H4O3 — ПВК (пируват, пировиноградная кислота) восстанавливается до молочной кислоты C3H4O3.
Гликолиз характерен для всех живых организмов и является древнейшим способом получения энергии.
У низших животных молочная кислота в результате брожения может перейти в этанол (дрожжи), ацетон (некоторые прокариоты). У многоклеточных молочная кислота способна накапливаться при сильных и длительных нагрузках.
b. Брожение
Распространено широко.
Большинство веществ поддается сбраживанию.
Конечные продукты брожения — этанол, ацетон, масляная кислота, бутанол.
Образование АТФ малоэффективно, т.к. образуются соединения со многими ковалентными связями. - Кислородный (дыхание)
Окисление протекает в митохондриях.
Конечными продуктами расщепления являются CO2 и H2O:
C6H12O6 (2C3H6O3)+ 6O2 + 36Ф + 36АДФ → 6CO2 + 6H2O + 36АТФ
- Кислородный этап в 18 раз эффективнее бескислородного.
- Учитывая бескислородный и кислородный этапы в результате расщепления одной молекулы глюкозы образуется 38 молекул АТФ.
В митохондриях кроме глюкозы расщепляются жирные кислоты.
Типы питания
По типу питания все живые организмы делятся на две группы:
- Автотрофы («сам себя питает») — фототрофы и хемотрофы
- Гетеротрофы — сапрофиты и паразиты
- Источник органического вещества CO2, H2O, минеральные вещества.
- Источник энергии — солнце.
- Представители — эвглена зеленая, все виды растений, цианобактерии, оксифотобактерии.
- NH3, H2S, FeO, H2O.
- Экзотермические окислительно-восстановительные реакции.
- Нитробактерии, серобактерии, железобактерии.
- Мертвые тела.
- Экзотермические реакции окисления органических веществ.
- Большинство животных, грибы, бактерии.
- Органические вещества живых тел.
- Аналогично сапрофитам.
- Насекомые, круглые и плоские черви, простейшие, вирусы, фаги, грибы (микозы), растения (петров крест).
Существует ряд организмов питающихся автотрофно и гетеротрофно (подкласс растительных жгутиконосцев — эвглена зеленая).
Фотосинтез
В год растениями фиксируется примерно 170*10 6 тонн углерода.
В результате образуется примерно 40*106 тонн органических веществ.
КПД фотосинтеза около 1%.
Продуктивность растений = 1 г/м2*ч (грамм на квадратный метр в час).
КПД у одноклеточной водоросли хлореллы достигает 10%.
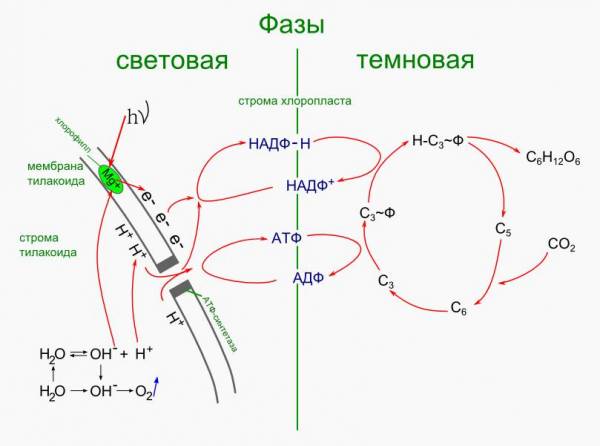
Фотосинтез в хлоропластах протекает в два этапа. В тилакоидах гран протекают реакции, вызванные светом (световые реакции). В строме (матриксе) хлоропласта протекают реакции, несвязанные со светом (темновые реакции).
- Свет (hν) попадает на молекулу хлорофилла, находящуюся в мембране теллакоида граны. Это вызывает потерю одного e — у Mg. Электроны накапливаются на наружной стороне мембраны, создавая отрицательный заряд.
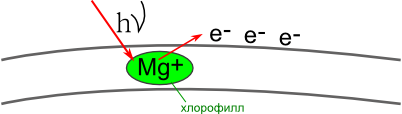
- Место ушедшего электрона занимает e — воды.
Реакция фотолиза:
H2O ↔ H + + OH — (катализатор hν)
OH — — 1e — → OH
4OH → 2H2O + O2 - Протоны водорода накапливаются на внутренней стороне мембраны, создавая положительный заряд.
Между внутренней и наружной сторонами создается разность потенциалов. - При достижении критического уровня разности потенциалов, что равно 200мВ (милливольт), протоны водорода устремляются наружу по протонному каналу фермента АТФ-синтетазы.
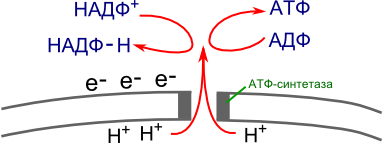
Полученная при этом энергия идет на синтез АТФ. - Водород на внешней стороне мембраны соединяется с e — и восстанавливает фермент НАДФ (никотинаминоадениндинуклиотидфосфат):
НАДФ + (окисленная форма) + 2e — + H + → НАДФ-H (восстановленная форма)
В световые реакции поступают hν, H2O, АДФ и НАДФ + . В результате образуются O2, АТФ, НАДФ-H.
АТФ и восстановленный НАДФ поступают в темновые реакции.
Темновые реакции (цикл Кальвина):
Открыл Кальвин в 1962 г.
Темновые реакции проходят в строме хлоропласта, куда поступают АТФ, восстановленный НАДФ-H, CO2. Кроме этого в строме постоянно находятся пентозы — C5H10O5.
- К пентозе присоединяется CO2, при этом образуется неустойчивая гексоза:
C5. + CO2 → C6.
, которая сразу же распадается на две триозы:
C6. → 2C3. - Триозы насыщаются от АТФ энергией в виде фосфорного остатка с макроэргической связью:
C3. + АТФ → C3.
Ф + АДФ
Далее триозы восстанавливается НАДФ-Н (насыщаются водородом):
C3.
Ф + НАДФ+
a. Из двух триоз образуется гексоза (глюкоза), которая уходит из хлоропласта:
2H-C3.
Ф → C6H12O6
b. Часть триоз объединяется в пентозы, которые возвращаются в цикл Кальвина:
5C3. → 5C5.
Схема световой и темновой фаз фотосинтеза: